
Section 12.5: Zinc as Lewis Acid and Template
- Page ID
- 440908
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
Introduction
Unlike most other biologically important metals, zinc is not redox active and does not directly participate in catalytic reaction. In biological systems zinc is primarily found as Zn2+, which is a d10 ion, making it unfavorable to either oxidize or reduce. Zinc ions do play important biochemical roles. They can act as Lewis acids, to indirectly catalyse reactions such as the hydration of carbon dioxide in carbonic anhydrase. They can also act as templates or scaffolds for protein tertiary structure.
Zinc in Carbonic Anhydrase
Chemical Reaction
Carbon-dioxide (CO2) hydration and its mechanism in living systems are of fundamental importance for bioinorganic chemistry. In 1932 the existence of an enzyme catalyzing CO2 hydration in red blood cells, carbonic anhydrase, was established. In 1939 the enzyme was recognized to contain zinc. Because CO2 is both the starting point for photosynthesis and the endpoint of substrate oxidation, carbonic anhydrases are now known to be ubiquitous, occurring in animals, plants, bacteria, and fungi. Carbonic anhydrase is a classic example of a hydrolytic enzyme, one that catalyzes addition or removal of water to a substrate molecule. More specifically, it catalyzes the reversible conversion of carbon dioxide (CO2) to bicarbonate (HCO3-).

Although hydration of CO2 is spontaneous in water at pH 7, the reaction is kinetically slow (k = 10-1 s-1), too slow to convert all CO2 produced in respiration. Only above pH 9 does the uncatalyzed reaction become fast, due to direct attack of OH-, which is a much better nucleophile than H2O (k = 104 M-1 s-1, in the presence of OH-). Figure \(\PageIndex{2}\) compares nucleophilic attack of water versus hydroxide (OH-) on CO2.
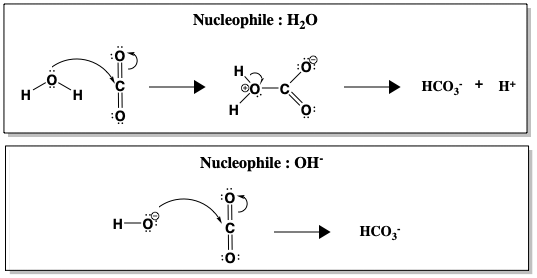
Figure \(\PageIndex{2}\): Nucleophilic attack on carbon in carbon dioxide (CO2) in the formation of bicarbonate (HCO3-). In the top panel, H2O is the nucleophile. The bottom panel shows the reaction with hydroxide (OH-) as the nucleophile. Since the OH- is a stronger nucleophile, HCO3- formation is faster when OH- is the nucleophile.
Between H2O and OH-, formation of HCO3- occurs faster when OH- is the nucleophile. A faster reaction at higher pH, when more OH- ions are present, suggests OH- is involved in the rate determining step. However, realistically, the pH of human blood cannot be changed to speed up hydration of CO2. Instead, humans use carbonic anhydrase to catalyse the reaction. When carbonic anhydrase is present, the reaction is sped up to a rate of k = 106 s-1.
The ubiquity of carbonic anhydrase in different organisms reflects the importance of these enzymes in sustaining life. The speed of catalyzed CO2 hydration is essential to meet the needs of living cells. Some physiological carbonic anhydrase functions include pH regulation, electrolyte secretion, ion transport, and CO2 homeostasis. In the digestive tract, carbonic anhydrases plays a role in the secretion of acid and keep saliva neutral by modulating pH. Among these functions, CA most notably plays a role in transport of CO2 and HCO3- related to respiration. With low blood solubility, CO2 must be converted to a more soluble form, HCO3-, for transport throughout the body. Bicarbonate ions eventually reach the lungs, get converted back to CO2, and exit the body through exhalation.
Medical research revolving around carbonic anhydrase focuses on the Zn-containing active site as a therapeutic target for various disease treatments; both carbonic anhydrase inhibitors and activators are incorporated in drug design. Carbonic anhydrase inhibitors are used as treatment for epilepsy, ulcers, cancer, obesity, and other neurological disorders. In the eye, carbonic anhydrase produces hydrogen ions that maintain optic pressure. However, too much pressure in the eye can damage the optic nerve and cause glaucoma. Carbonic anhydrase activity can create a concentration gradient that drives the transport of water to the optical nerve. When too much water is around the optical nerve, pressure around the nerve increases causing damage. Inhibition of carbonic anhydrase has become a key treatment of glaucoma.
Role of Zinc
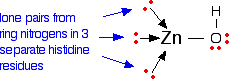
The zinc ion is bound to the protein via three links to separate histidine residues in the chain, shown in pink in Figure \(\PageIndex{3}\). The zinc is also attached to a hydroxide ion shown in the picture using red for the oxygen and white for the hydrogen.

If you look at the model of the arrangement around the zinc ion in the picture above the arrangement of the four groups around the zinc is approximately tetrahedral.
How does this catalyse the reaction between carbon dioxide and water?
A carbon dioxide molecule is held by a nearby part of the active site so that one of the lone pairs on the oxygen is orientated towards the carbon atom of the carbon dioxide molecule. Binding the carbon dioxide in the enzyme also increases the existing polarity of the carbon-oxygen bonds.
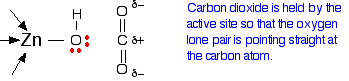
If you have done any work on organic reaction mechanisms at all, then it is pretty obvious what is going to happen. The lone pair forms a bond with the carbon atom and part of one of the carbon-oxygen bonds breaks and leaves the oxygen atom with a negative charge on it. This results in a hydrogen carbonate ion coordinated to the zinc center.
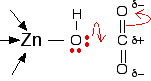
The next step in the reaction is for the hydrogen carbonate to leave and be replaced with a water molecule from the cell solution.
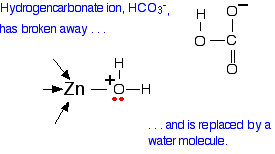
All that now needs to happen to get the catalyst back to where it started is for the water to lose a hydrogen ion. This is transferred away from the zinc center and eventually out of the active site completely through the network of ordered, hydrogen bonded water molecules shown in Figure \(\PageIndex{4}\). The carbonic anhydrase enzyme can do this sequence of reactions about a million times a second.
Zinc as a Structural Template
Many proteins use Zn2+ as a structural scaffold for protein folding. These proteins employ "zinc finger" domains that bind to Zn through combinations of cysteine (Cys) and Histidine (His) side chains. Zinc binding to these domains stabilizes a specific protein fold that is critical to the protein's function. Over half of transcription factors have zinc finger domains that allow the proteins to recognize and bind to DNA. Many other proteins also have zinc-finger domains and these domains are important for a variety of functions including DNA-binding, RNA-binding, protein-protein interactions, and catalysis.
The human glioblastoma protein (GLI) family is a class of transcription factors that demonstrate sequence-specific DNA binding through "classic" Cys2His2 Zn-binding motifs. These proteins are essential during embryonic development. Dysfunction of these proteins are linked closely to glioblastoma and other aggressive cancers. The zinc finger domain of GLI1 is shown in this image (PDB 2gli),2 where the protein surface is highlighted in yellow. GLI1 possesses five Cys2His2 zinc finger domains. When Zn is not bound to the protein, it does not have the appropriate structure to bind to recognize DNA.

The size and structure of the Zn2+ ion is critical to provide the correct protein fold for DNA binding. Thus, zinc fingers must bind specifically to Zn (as opposed to other biologically available metal ions). The zinc finger domains use principles of inorganic chemistry to control metal ion selectivity. While other metal ions, like Co2+, Ni2+, Fe2+, and Mn2+, can bind in the zinc finger sites, Zn2+ binds 1,000 to 100,000 times more strongly than any of these other metals.
Exercise \(\PageIndex{1}\)
Apply HSAB theory to explain why there is such a strong selectivity for Zn2+ binding in a zinc finger site as opposed to other biologically available metals, including Co2+, Ni2+, Fe2+, and Mn2+.
- Answer
-
The zinc binding site contains two borderline histadine ligands and two soft cystine ligands. The Zn2+ is a softer borderline Lewis acid compared to other biologically available metal ions, especially Fe2+. This is because the size of transition metal cations don't vary much across the period, giving them all similar size to charge ratios and Zn2+has the highest number of valence electrons, making it the most polarizable. Because the active site has two soft base donors and two borderline base donors, softer metals will preferentially bind in the active site.
Exercise \(\PageIndex{2}\)
Cadmium (Cd2+) and lead (Pb2+) are toxic metal ions that bind more strongly than Zn2+ to zinc binding sites in proteins. Why do these toxic metals compete well with Zn2+, while other metals like Co2+, Ni2+, Fe2+, and Mn2+ don’t?
- Answer
-
As discussed in Exercise 1, the zinc finger binding site is biased towards coordinating softer metals. Both cadmium and lead ions are much softer than zinc and will have a stronger affinity than zinc for the zinc binding site. Under normal conditions this is not an issue because lead and cadmium are not present in significant concentrations.
Exercise \(\PageIndex{1}\)
If a toxic metal, like Cd2+ would bind to the Zn2+ binding site, how and why might this affect the protein’s ability to bind to DNA?.
- Answer
-
The proper folding and structure of these proteins is dependent on zinc coordination in the zinc finger domains. If a larger metal ion, such as Cd2+ were to bind in place of Zn2+ this would change the size and/or geometry of the zinc finger domain and therefore the entire protein. The correct shape of the protein is essential for recognition and binding of DNA.
References
-
Inspired by Zinc Finger Proteins: A Bioinorganic Example of LFSE and HSAB, a problem set created by Elizabeth R. Jamieson, Smith College (ejamieso@smith.edu) and posted on VIPEr (www.ionicviper.org) on June 24, 2013. Copyright Elizabeth R. Jamieson 2013. This work is licensed under the Creative Commons Attribution-NonCommerical-ShareAlike 3.0 Unported License. To view a copy of this license visit http://creativecommons.org/about/license/.
-
Pavletich, N. P.; Pabo, C. O., Crystal Structure of a Five-Finger Gli-DNA Complex: New Perspectives on Zinc Fingers. Science 1993, 261 (5129), 1701-7.
-
Munro, D.; Ghersi, D.; Singh, M., Two Critical Positions in Zinc Finger Domains Are Heavily Mutated in Three Human Cancer Types. PLOS Computational Biology 2018, 14 (6), e1006290.