
13.3: An Overview of Fatty Acid Metabolism
- Page ID
- 106373

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
In the introduction to this chapter, we learned about a patient suffering from a rare disease affecting fatty acid metabolism. The reaction mechanisms that we are about to learn about in the next two sections are central to the process by which fatty acids are assembled (synthesis) and taken apart (degradation), so it is worth our time to go through a brief overview before diving into the chemical details.
Fatty acid metabolism is a two-carbon process: in the synthetic directions, two carbons are added at a time to a growing fatty acid chain, and in the degradative direction, two carbons are removed at a time. In each case, there is a four-step reaction cycle that gets repeated over and over. We will learn in this chapter about steps I and III in the synthesis direction, and steps II and IV in the degradative direction. The remaining reactions, and the roles played by the coenzymes involved, are the main topic of chapter 15.
'ACP' stands for 'acyl carrier protein', which is a protein that links to growing fatty acid chains through a thioester group (see section 11.5)
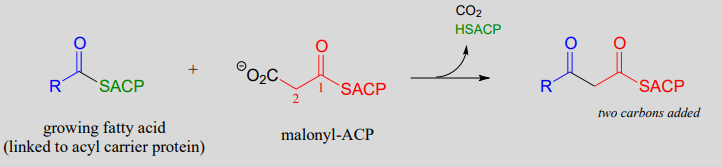
Step I: Condensation (covered in section 13.3)
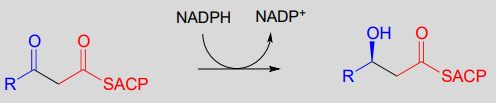
Step II: Ketone hydrogenation (covered in section 15.3)
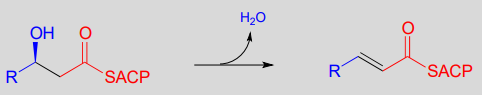
Step III: Elimination (covered in section 13.4)
Step IV: Alkene hydrogenation (covered in section 15.4)
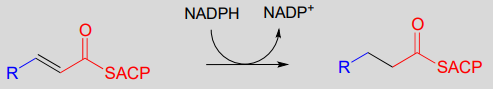
. . . back to step I, add another malonyl-ACP, repeat.
Step I: Alkane oxidation (covered in section 15.4)
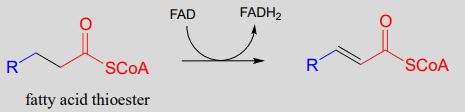
Step II: Addition of water (covered in section 13.4)
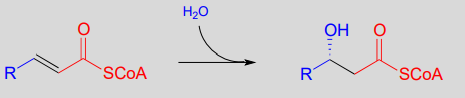
Step III: Oxidation of alcohol (covered in section 15.3)
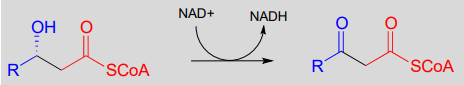
Step IV: Cleavage (covered in section 13.3)
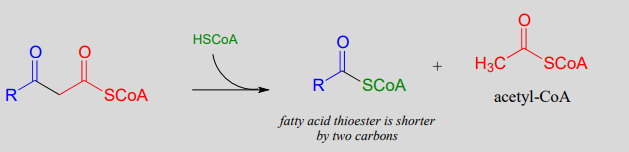
. . . back to step I
When looking at these two pathways, it is important to recognize that they are not the reverse of each other. Different coenzymes are in play, different thioesters are involved (coenzyme A in the degradative direction, acyl carrier protein in the synthetic direction), and even the stereochemistry is different (compare the alcohols in steps II/III of both pathways). As you will learn in more detail in a biochemistry course, metabolic pathways that work in opposite directions are generally not the exact reverse of each other. In some, like fatty acid biosynthesis, all of the steps are catalyzed by different enzymes in the synthetic and degradative directions. Other 'opposite direction' pathways, such as glycolysis/gluconeogenesis, contain mostly reversible reactions (each catalyzed by one enzyme working in both directions), and a few irreversible 'check points' where the reaction steps are different in the two directions. As you will learn when you study metabolism in biochemistry course, this has important implications in how two 'opposite direction' metabolic pathways can be regulated independently of one another.
Recall that in Chapter 12 we emphasized the importance of two reaction types - the aldol addition and the Claisen condensation - in their role in forming (and breaking) most of the carbon-carbon bonds in a living cell. We have already learned about the aldol addition, and its reverse, the retro-aldol cleavage. Now, we will study the Claisen condensation reaction, and its reverse, the retro-Claisen cleavage. Step I in fatty acid synthesis is a Claisen condensation, and step IV in fatty acid degradation is a retro-Claisen cleavage.
In section 13.4, we will look more closely at the reactions taking place in step III of fatty acid synthesis (an elimination) and step II of fatty acid degradation (a conjugate addition).