
Enzyme Inhibitors
- Page ID
- 3966

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)This page looks at the effect of inhibitors on reactions involving enzymes. This is the third and final page talking about how enzymes function as catalysts.
Competitive Inhibitors
This is the most straightforward and obvious form of enzyme inhibition - and the name tells you exactly what happens. The inhibitor has a similar shape to the usual substrate for the enzyme, and competes with it for the active site. However, once it is attached to the active site, nothing happens to it. It doesn't react - essentially, it just gets in the way. Remember the general equation for an enzyme reacting with a substrate?
\[\ce{E + X <=> E-S}\, \text{complex} \ce{-> E + P}\]
The equivalent equation for a competitive inhibitor looks like this:
\[ \ce{E + I_{c} <=> E-I_{c}}\, \text{complex}\]
The complex does not react any further to form products - but its formation is still reversible. It breaks up again to form the enzyme and the inhibitor molecule. That means that if you increase the concentration of the substrate, the substrate can out-compete the inhibitor, and so the normal reaction can take place at a reasonable rate.
A simple example of this involves malonate ions inhibiting the enzyme succinate dehydrogenase. This enzyme catalyses the conversion of succinate ions to fumarate ions. The modern names are:
- malonate: propanedioate
- succinate: butanedioate
- fumarate: trans-butenedioate
The conversion that succinic dehydrogenase carries out is:
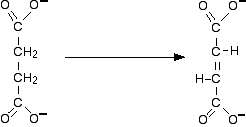
The reaction is inhibited by malonate ions which have a very similar shape to succinate ions.
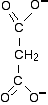
The similar shape lets the malonate ions bind to the active site, but the lack of the CH2-CH2 bond in the centre of the ion stops any further reaction taking place. The malonate ions therefore block the active site - but remember that this is reversible. The malonate ions will break away and free up the enzyme again. The malonate ions are in competition for the site - they aren't destroying it. If the succinate ions have a greater concentration than the malonate ions, by chance they will get access to the site more often than the malonate ions. That means that you can overcome the effect of a competitive inhibitor by increasing the concentration of the substrate.
Non-competitive inhibitors
A non-competitive inhibitor doesn't attach itself to the active site, but attaches somewhere else on the enzyme. By attaching somewhere else it affects the structure of the enzyme and so the way the enzyme works. Because there isn't any competition involved between the inhibitor and the substrate, increasing the substrate concentration won't help. You will find two explanations for this. We'll look at the simple, fairly obvious one in some detail in a minute. I want to have a brief word about the other one first.
"Pure" non-competitive inhibitors
This explanation says that the inhibitor doesn't affect the ability of the substrate to bond with the active site, but stops it reacting once it is there. I found a couple of biochemistry sites which said that inhibitors working like this (which they describe as pure non-competitive inhibitors) are virtually unknown. As a non-biochemist, I don't know what the truth is about this - if you want to find out, you will probably have to do a biochemistry degree!
Other non-competitive Inhibitors
The straightforward explanation (which would seem to apply to most enzymes) is that reaction with the inhibitor causes the shape of the active site to change. Remember that non-competitive inhibitors aren't attaching directly to the active site, but elsewhere on the enzyme. The inhibitor attachs to a side group in the protein chain, and affects the way the protein folds into its tertiary structure. That in turn changes the shape of the active site. If the shape of the active site changes, then the substrate can't attach to it any more. Some non-competitive inhibitors attach irreversibly to the enzyme, and therefore stop it working permanently. Others attach reversibly.
A relatively uncomplicated example of non-competitive inhibitors in a reasonably familiar situation is heavy metal poisoning. You are probably aware that compounds containing heavy metals such as lead, mercury, copper or silver are poisonous. This is because ions of these metals are non-competitive inhibitors for several enzymes. I'm going to take silver as a simple example. Silver ions react with -SH groups in the side groups of cysteine residues in the protein chain:

There is not enough electronegativity difference between silver and sulfur for a full ionic bond and so the bond can be considered as covalent. If the cysteine residue is somewhere on the protein chain which affects the way it folds into its tertiary structure, then altering this group could have an effect on the shape of the active site, and so stop the enzyme from working. The 2+ ions from, for example, mercury, copper or lead can behave similarly - also attaching themselves to the sulphur in place of the hydrogen.