
Glutamine Synthetase
- Page ID
- 98114

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
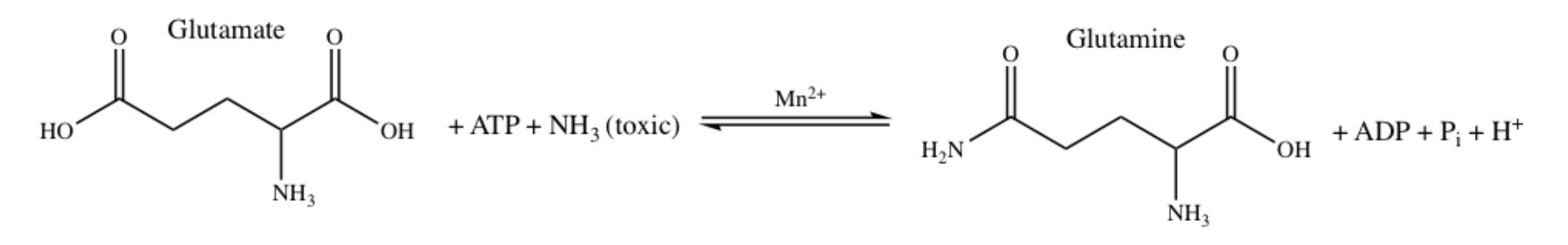
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Glutamine synthetase (GS) is an enzyme that is responsible for catalyzing the reaction that synthesizes glutamine from glutamate and ammonia, demonstrated in Figure 1 below.1 Additionally, GS catalyzes the reverse reaction of glutamate to glutamine, the “7-glutamyl transfer reaction” which transfers the 7-glutamyl from glutamine to hydroxylamine yielding 7-glutamyl hydroxamate, and more.16 This paper will focus on the synthesis of glutamate to glutamine.
Ultimately, GS regulates the levels of nitrogen in molecules such as DNA and amino acids; it takes the nitrogen from the glutamine produced in the synthesis reaction and will deliver it to nitrogen-deficient molecules. GS could also slow the transfer of nitrogen to nitrogen-rich molecules, as well as regulate the production of certain amino acids and nucleotides, like adenosine monophosphate (AMP), histidine, and alanine, by reducing the synthesis of these molecules when they are in excess.2 GS is present in both plants and animals. In plants, GS detoxifies ammonia, and in animals it recycles amino acid neurotransmitter molecules and detoxifies ammonia.3
Human GS is present in both the brain and the liver. In the human brain, GS is most often found in astrocytes, star-shaped glial cells in the nervous system. Astrocytes are responsible for protecting neurons against excitotoxicity (damage or death of nerve cells by stimulation from neurotransmitters). This protection is accomplished through the conversion of excess ammonia, which is toxic, and glutamate, which is neurotoxic, into glutamine, catalyzed by GS.4 In both the brain and the liver, the role of human GS is similar to bacterial GS in that the human GS eliminates toxic ammonia. Malfunction of this enzyme in the liver can lead to liver cirrhosis, chronic hepatitis B and C, some types of hyperplasia and some neoplasms. As for human GS in the brain, it monitors the toxicity of glutamate by converting glutamate into glutamine, which is less neurotoxic (Figure 1).15 Malfunction of GS in the brain can lead to neurological diseases such as Alzheimer’s, epilepsy, glioblastoma multiforme, anxiety and depression.11
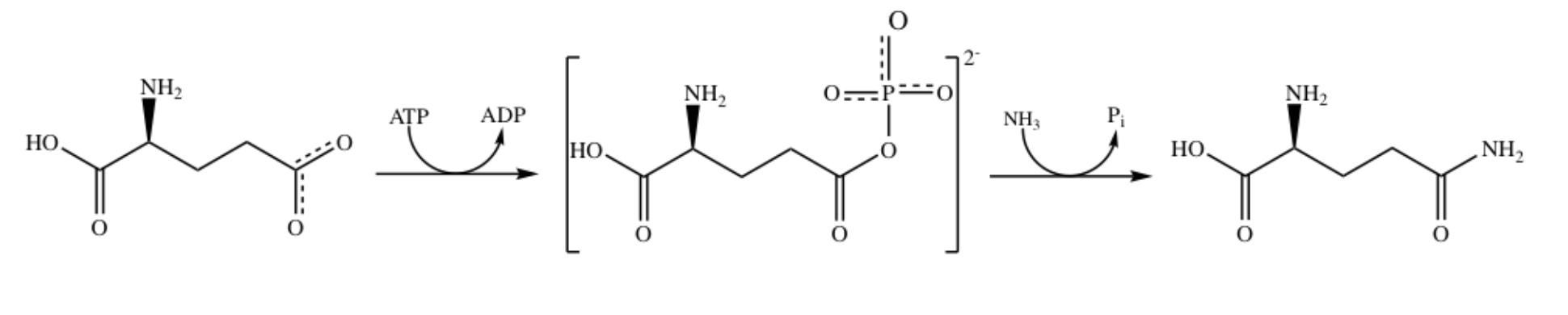
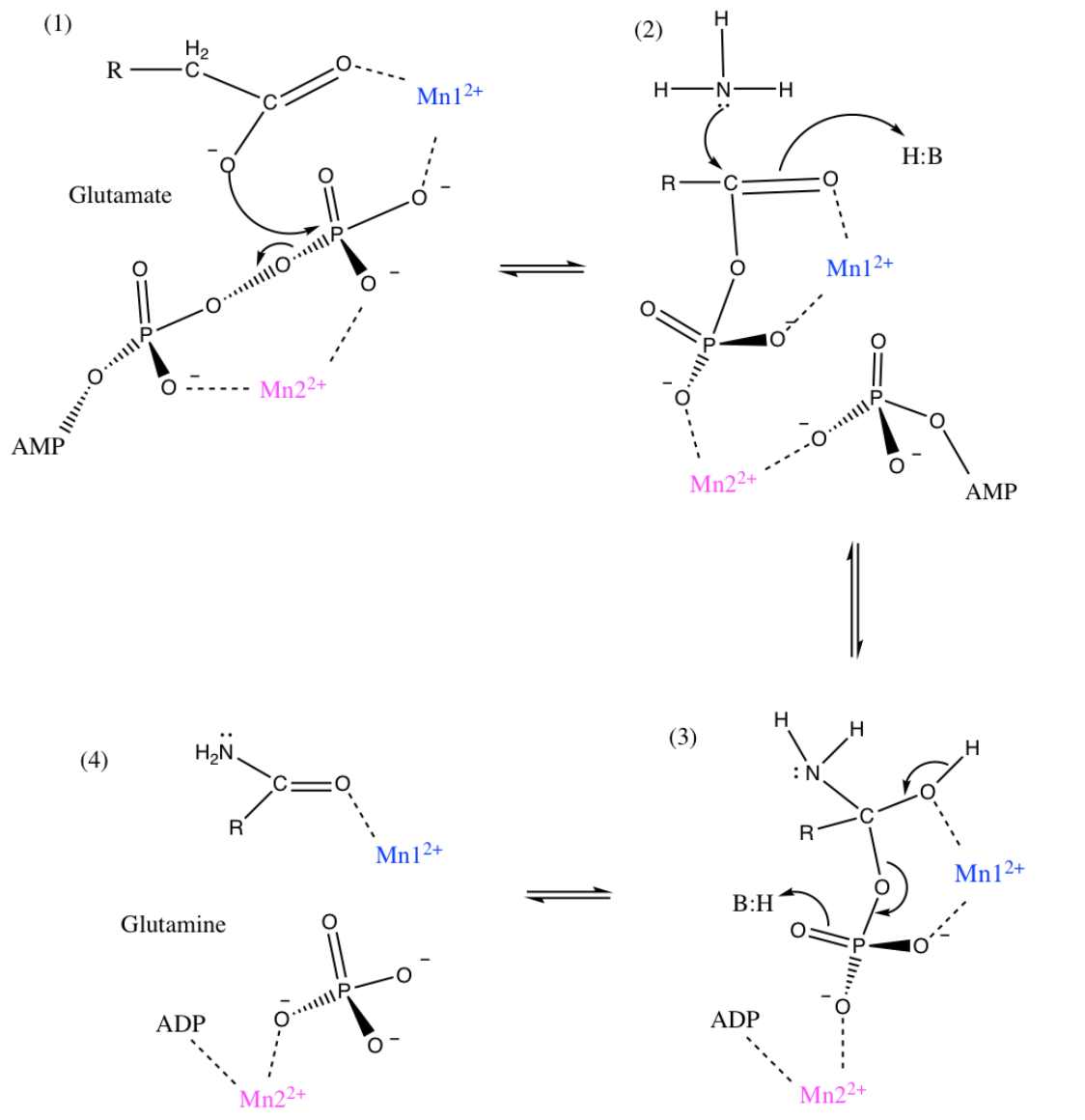
While Figure 1 demonstrates the overall reaction of glutamate to form glutamine, it is proposed that this reaction occurs in two steps, with a tetrahedral intermediate. The proposed two step mechanism is shown in Figure 2 below.
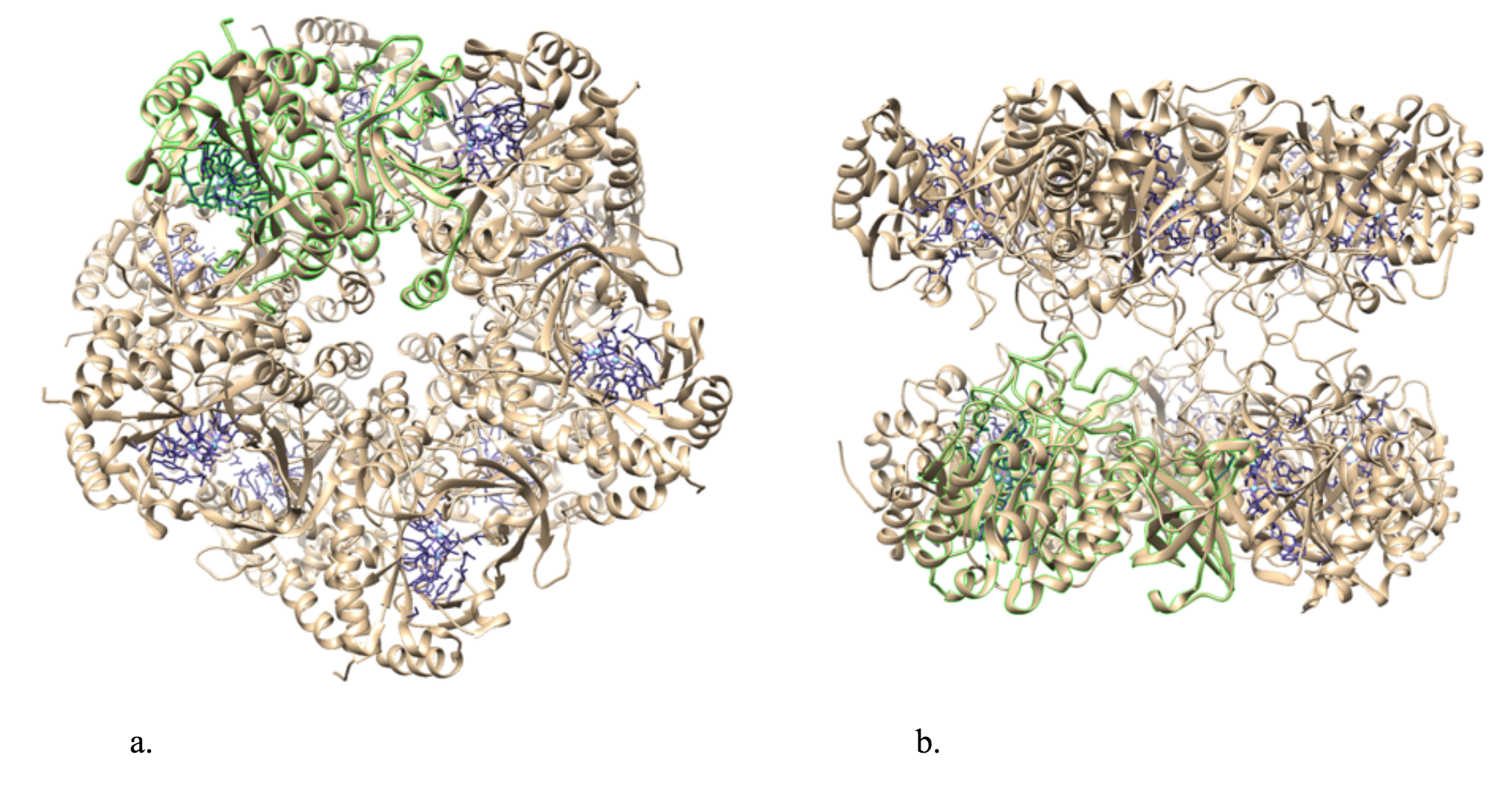
The general structure of bacterial GS is a dodecamer formed from two face-to-face hexameric rings of subunits. There are twelve active sites, one in each monomer.5 Figure 3 below illustrates the structure of bacterial glutamine synthetase.

Each active site can be described as a bifunnel in which ATP binds at the top and glutamate binds at the bottom. Two metal cations lie at the center of each active site. The enzyme is functional with either Mg2+ or Mn2+ in the active site, however Mn2+ is more common despite the fact that it slightly slows down the activity of the GS enzyme.5 GS catalysis is faster with Mg2+, however GS selects Mn2+ because it binds to the enzyme with higher affinity.7 Figure 4 below shows the structure of a single active site of GS. There are multiple ligands bound in the active sites of GS. The pink Mn2+ metal ion illustrated below binds to two glutamate amino acid side chains, a histidine side chain and two water molecules. The blue Mn2+ metal ion illustrated below binds to three glutamate amino acid side chains and two water molecules. Figure 4 also illustrates that the pink Mn2+ binds to an adenosine diphosphate (ADP) molecule, which is a common substrate for GS, and that the blue metal ion binds to a phosphinothricin (PPT) molecule, which is an inhibitor of GS that binds to the enzyme in a similar position as the amino acid substrate (glutamate or glutamine).14 This demonstrates that, while PPT and ADP are not always bound in the active site, the metal ions in GS are capable of binding to six ligands each. Ultimately, GS is geometrically flexible, allowing substrates to be both bound and released in/from the active sites; catalytic function is dependent on its structure. This active site would not be able to function if the metal ion was coordinately saturated, meaning that all of the coordination sites are occupied. The system must have one exchangeable coordination position so that the substrate ligand can interact with the metal, react, and then allow for product to be released or exchanged for new substrate. It is known that GS has three distinct substrate binding sites: one for nucleotide, one for ammonium ion and one for amino acids.5 It binds its substrates near its two metal ion sites, however, the order of binding remains unclear.16 The amino acid substrate binding is very specific in bacterial and eukaryotic GS, whereas the nucleotide binding site varies much more in what it can bind. This lead to the determination that the nucleotide binding sites could be used for target drug design.15

While this paper will focus on bacterial GS and how it functions, it is also interesting to note that there is another form of GS found in humans. The structure of human GS varies slightly from bacterial GS. It consists of a homodecamer structure organized in two pentameric rings (as opposed to hexameric rings) in which the active site is still between adjacent monomers, forming a bifunnel. This is illustrated in Figures 5 and 6 below. It is also different from the bacterial GS in that there are three metal ions in the active site, instead of two.15 While at first it was thought that the functional and structural properties of the active site of GS were conserved in mammals and plants, later studies found crystal structures of human GS actually contained three metal ions in each active site.11,15
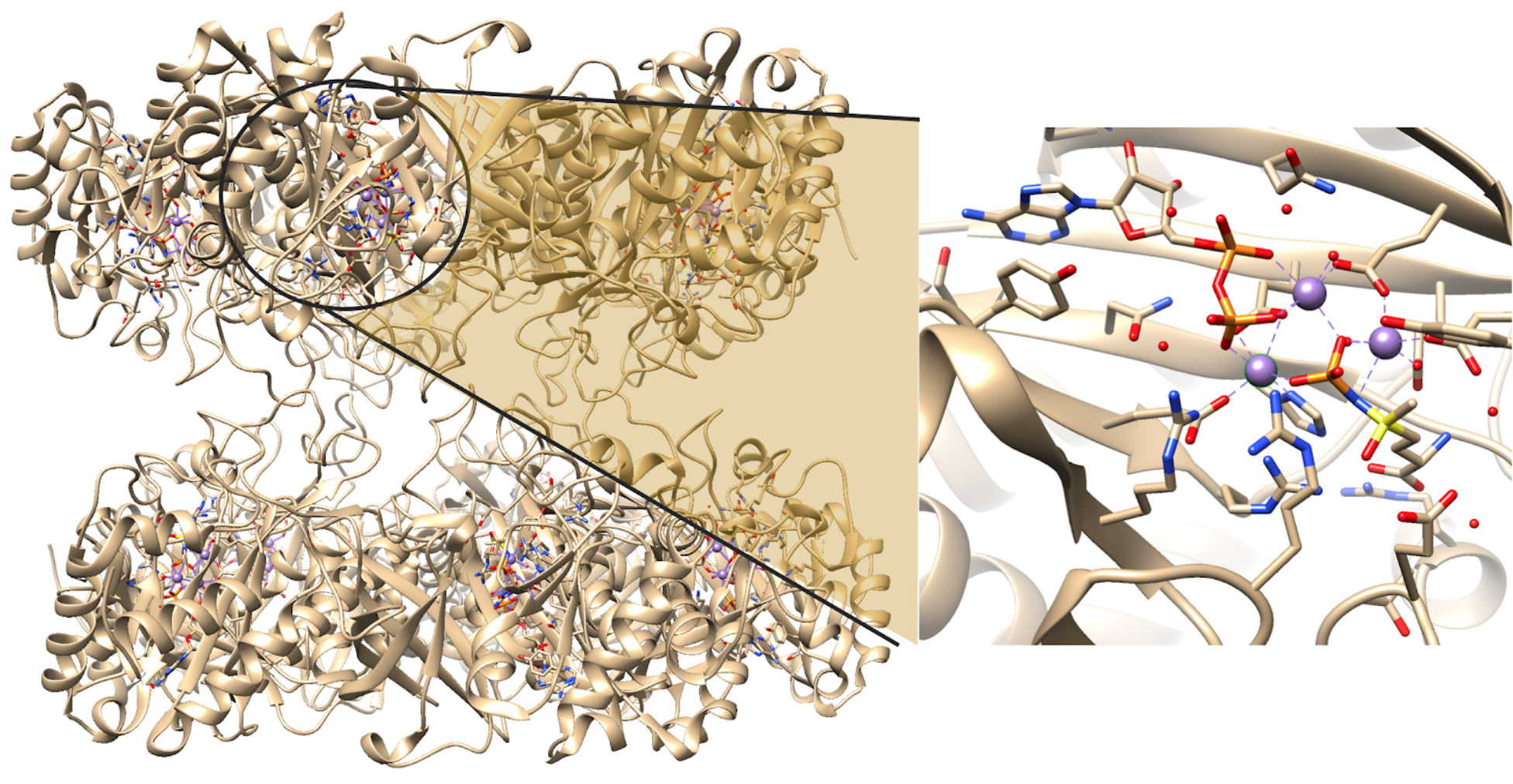
As stated, there are two metal ions in the active sites of bacterial GS, one binds with high affinity and the other with low.2 The high affinity metal binding site acts as the catalytic cofactor, represented in blue below, which means that it is a non-protein metallic ion that is required for an enzyme's activity. The metal ion (Mn2+) stabilizes both the electrostatic interactions and Lewis acid catalysis, providing the active-site structure and stabilization of intermediates for the reaction (Figure 7).1 The low affinity metal ion aids in the stabilization of the active site of GS and with binding of the Mn-ATP chelate.1,5 The overall reaction for the synthesis of glutamine from glutamate through the GS catalyst is shown above (Figure 1).5 This reaction is important for the uptake of nitrogen by organic compounds because glutamine serves as a building block for the biosynthesis of various metabolites.8 The mechanism for the conversion of glutamate to glutamine is demonstrated below in Figure 7.
Given the role of these two metal ions, it is important to know more about them to gain a better understanding as to how GS functions. Both metal ions have an oxidation state of +2. Most often, Mn2+ is the metal ion because GS has a much higher binding affinity for Mn2+ (300 - 1000x) than it does for magnesium (Mg2+), which has the second highest affinity.5 Due to its +2 charge, the manganese ion (atomic number 25) has five d-electrons.
The selective binding of Mn2+ to GS can be explained through several theories. While other metal ions, including Cd2+, Mg2+, Ca2+, Co2+, and Zn2+, were tested for their affinity to GS, Mn2+ was found to be the most effective metal ion, second to Mg2+. With respect to the Hard-Soft-Acid-Base theory (HSAB), it makes sense that GS binds to Mn2+ ions. HSAB categorizes metals (acids) and ligands (bases) by their charge densities, polarizability, and electrostatic or covalent bonding character. The harder something is, the less polarizable it is. Generally, harder atoms are associated with electrostatic interactions, which includes the alkaline and alkali earth metals, whereas softer atoms are associated more with polarizability and covalently bonding to ligands. There is a general rule to follow: hard acids prefer to bind to hard bases and soft acids prefer to bind to soft bases. Mn2+, Mg2+, Ca2+ are classified as a hard acids. However, Cd2+ is a soft acid and Co2+ and Zn2+ are borderline acids. It is predicted that the metal ions bind to water, to a carboxylate oxygen on glutamate, and to the imidazole nitrogen on histidine. Oxygen donor groups are classified as hard bases, while the imidazole N is borderline. Thus, this binding site has mixed hard/borderline character, with most donor groups contributing hard oxygen ligands. Considering the binding site character, we can apply HSAB theory to predict that hard metal ions Mn2+, Mg2+, Ca2+, and Co2+ would be prefered in this GS cation active site over soft and borderline metals like Cd2+, and Zn2+.
GS requires a metal ion that is geometrically flexible so that substrates can be both bound and broken from the active site. The metal ion should be capable of adopting three to five coordinate geometries. Co2+ is a d7 metal ion meaning that it has seven electrons in its shell. Considering the 18-electron rule, it would need five or six ligands to be coordinatively saturated with about eighteen electrons (actually seventeen or nineteen because of the odd number of d-electrons). Additionally, LFSE causes Co2+ to have a preference for six-coordinate octahedral geometry. Ligand Field Stabilization (LFSE) is defined as the total energy of the electrons relative to the average energy of the orbitals. For octahedral complexes, LFSE depends on the number of electrons in the lower energy orbitals (t2g) and the electrons in the higher energy orbitals (eg) relative to the barycenter (center of mass of two or more bodies that are orbiting each other). LFSE of an octahedral complex can be calculated with the following equation:
(1)
Where x is the number of electrons in the t2g orbitals and y is the number of electrons in the eg orbitals.
On the contrary, Ca2+ has zero d-electrons (d0), Mg2+ has zero d-electrons (d0), and Mn2+ has five d-electrons (d5). These metal ions are more geometrically flexible because they all have an LFSE of zero, calculated using Equation 1. With no stabilization by LFSE, these ions do not have a preference for a certain geometry. Thus, they would be content with binding to the three to five coordinate geometries that are available in the active site of GS. Because Co2+ would prefer an octahedral geometry as opposed to one with only three to five coordinate geometries, it is not in ideal metal ion for GS. This leaves three possible metal ions: Ca2+, Mg2+, and Mg2+. Another factor that may influence what metal ion is preferred by GS is the atomic size of the ions. Ca2+ has an ionic radius of 100 pm, whereas Mg2+ has an ionic radius of 72 pm and Mn2+ has an ionic radius of 70 pm.13 This explains why Ca2+ is not the preferred metal ion; it has a much larger ionic radius than that of Mg2+ and Mn2+. The smaller metal ions are preferred over the larger Ca2+. This leaves two remaining metal ions, Mg2+ and Mn2+. To explain why Mn2+ is the preferred metal ion to GS as opposed to Mg2+, we consider the effective nuclear charge (Zeff) in each case. In general, Zeff increases across the rows of a periodic table and down the columns. Thus, Mn2+ has a greater Zeff than does Mg2+, meaning that Mn2+, with a greater effective charge, is more attracted to the electron density of ligands than Mg2+. This is related to the well-known Irving-Williams series. Ultimately, it makes sense that Mn2+ is the preferred metal ion bound to the active site of glutamine synthetase.
Although the geometry of the manganese metal ion in GS is flexible, when it is in its octahedral coordination, the system would most likely be high spin. The high spin state fills each unoccupied orbital with a single electron before any orbitals are filled with two electrons; the lowest energy arrangement takes priority. Although it is not completely clear, for the purpose of this discussion, we assume that Mn2+ has an octahedral geometry. There are two ways that the field splitting diagram for Mn2+, in the octahedral configuration, could be drawn: one for high spin and one for low spin. Both of these are depicted in Figure 8 below. Since Mn2+ is a 3d5 metal with a +2 charge and has water as a ligand it should be high spin. Additionally, glutamate amino acids are pi donors which encourages small delta (the gap in energy between the t2g and eg in an octahedral complex) and high spin. Histidine may be a weak pi acceptor, which usually encourages low spin, however, this would not increase delta enough to favor the low spin. It is important to note that a phosphinothricin (PPT) molecule is bound to the low affinity manganese ion presented in Figure 3 above. This is an inhibitor that was experimentally introduced into the system, therefore, it is not always present in the system. When PPT is bound to the low affinity metal ion (pink n1 Mn2+), then this Mn2+ ion would have an octahedral geometry. When PPT is not bound to the metal ion, then there is an open coordination site for either a water or substrate molecules to bind to.
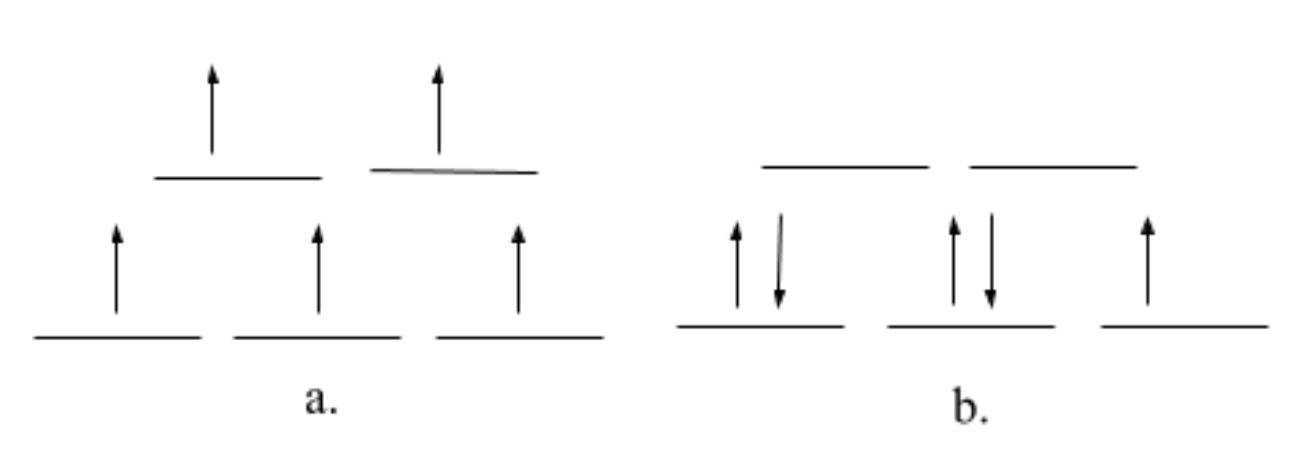
When it comes to kinetics, lability (or reactivity) refers to the rate with which ligands are replaced in coordination complexes; how quickly metal-ligand bonds are broken. There are many factors that influence whether a compound is labile or inert, including d-electron configuration, charge, LFSE, type of ligand, metal ionic radius and antibonding orbital occupancy. The more LFSE, the less labile the ion is (the less it will want to react). This makes sense, because the higher the ligand field stabilization energy there is, the less labile the metal (because it is more stable), thus, the molecule is more inert. Since Mn2+ has an LFSE of zero and the eg orbitals (antibonding orbitals) are occupied by electrons (high spin configuration), it is considered to be a kinetically labile metal ion which, again, is advantageous for the ligand/substrate binding and release in GS. In general, metals with zero LFSE are considered the most labile.
Glutamine synthetase is a multidentate chelator (a ligand that binds to a metal with more than one atom). The chelate effect is considered to be entropic, meaning that it increases the entropy (disorder) of a coordination reaction and thus increases the gibbs free energy. The chelate effect occurs upon binding of a metal ion to a multidentate ligand; the more atoms that the ligand uses to bind to the metal, the more stable the resulting complex. GS is a multidentate system because it provides more than two donor groups for the Mn2+ metal ions to bind to within the active site. More specifically, it is a tridentate chelator, providing three amino acid side chain ligands to each of the metal ions. These donor groups are on the protein side chains of glutamate, histidine and water molecules. This would help to explain why GS has a high affinity for a metal ion that can have multiple coordination sites.
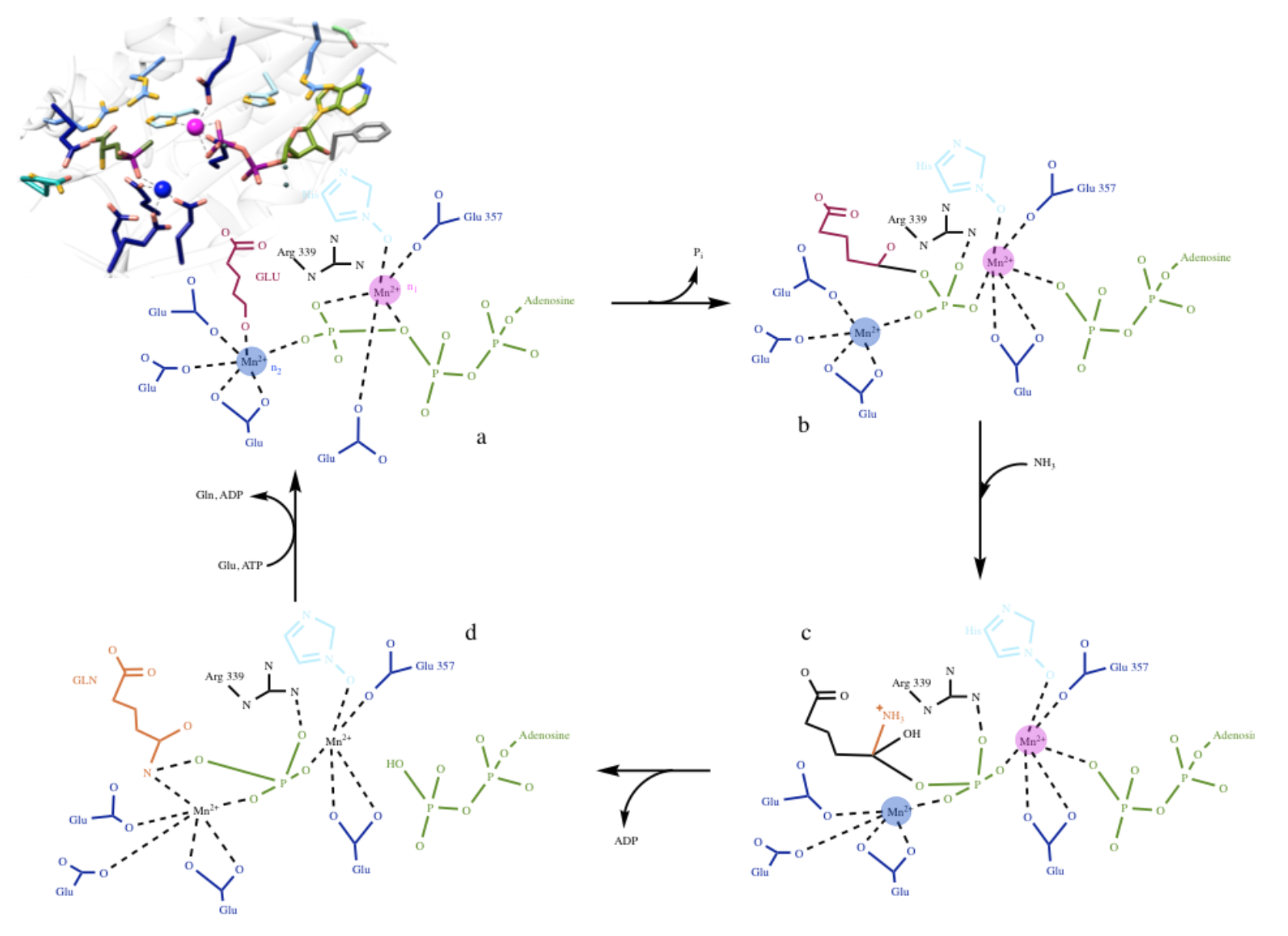
This brings us to the catalytic reaction. As stated previously, the geometry of each metal ion is thought to be pseudo octahedral. Each Mn2+ ion is bound to three amino acid side chains and two water molecules, in which one of the water molecules is shared between the two metal ions. It is known that the protein ligands of Mn2+ (the catalytic cofactor) are three glutamate amino acids. As for the second manganese ion, with lower binding affinity, it also has three amino acid side chain ligands, two glutamate amino acids and a histidine amino acid.9 The five coordinate pseudo-octahedral geometry leaves the metal ion with one open coordination site that can accommodate ligand binding. The proposed coordination geometry and mechanism is depicted in Figure 9 below.
When a complex absorbs light with an energy corresponding to delta, an electron is promoted from one d orbital to another, which is referred to as a d-d transition. Mn2+ has a partially-filled d-orbital, d5. Therefore, d-d transitions, or the excitation of electrons within the “d” orbitals (eg. From t2g to eg), can occur, which would result in possible electronic absorptions that can be observed with UV-vis spectroscopy. Because Mn2+ is a high spin metal ion, it has a small delta which means that it is easier to move electrons from the t2g to the eg. A small delta is an indication of low energy absorbing light. There could also be charge-transfer electronic transitions in this metal complex because, while there are not pi acceptor ligands, there are pi donor ligands. This means that charge transfers can occur from ligand to metal (LMCT). Studies have shown that the maximum Mn2+ absorbance value is when the detection wavelength at 553 nm.17 If Mn2+ absorbs at 553 nm, then that means it appears reddish-purple (magenta).
Currently, there are many active studies searching for GS inhibitors. A plant GS inhibitor (glufosinate) is one of the most widely used herbicides across the planet. Additionally, bacterial GS has been shown to be applicable in the development of drugs against tuberculosis as well as cancer therapy.11,12 There is still so much to learn about GS, including information about its structure and geometry, its metal ion binding properties and its catalytic and inhibition mechanisms.11
Sources
- Bertini, Gray, Stiefel, & Valentine. (2007). Biological Inorganic
- Chemistry. University Science Books.
- Goodsell, D. S. Glutamine Synthetase. RCSB Protein Data Bank 2002.
- Liaw, S., Kuo, I., & Eisenberg, D. (1995). Discovery of the ammonium substrate site on
- glutamine synthetase, a third cation binding site. Protein Science,4, 2358-2365. Retrieved March 4, 2018, from https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2143006/pdf/8563633.pdf.
- Suarez, I., Bodega, G., & Fernandez, B. (2002). Glutamine synthetase in brain: effect of ammonia. Neurochemistry International,41(2-3), 123-142. doi:10.1016/S0197-0186(02)00033-5
- Eisenberg, D.; Gill, H. S.; Pfluegl, G. M. U.; Rotstein, S. H. Structure–function Relationships of Glutamine Synthetases. Biochimica et Biophysica Acta (BBA) - Protein Structure and Molecular Enzymology 2000, 1477 (1), 122–145.
- Hunt, J. B.; Smyrniotis, P. Z.; Ginsburg, A.; Stadtman, E. R. Metal Ion Requirement by Glutamine Synthetase of Escherichia Coli in Catalysis of γ-Glutamyl Transfer. Archives of Biochemistry and Biophysics 1975, 166 (1), 102–124.
- Goodsell, D. (2002, June). Glutamine Synthetase. Retrieved March 05, 2018, from http://pdb101.rcsb.org/motm/30
- Schaller, C. P. (2017, September 08). The Chelate Effect. Retrieved March 06, 2018, from https://chem.libretexts.org/LibreTex...Chelate_Effect
- Yamashita, M. M., Almassy, R. J., Janson, C. A., Cascio, D., & Eisenberg, D. (1989). Refined Atomic Model of Glutamine Synthetase at 3.5 A Resolution*. The Journal of Biological Chemistry,264(Oct 25), 17681-17690. Retrieved February 14,
- 2018, from http://www.jbc.org.proxy.library.nd....17681.full.pdf
- Hunt, J. B.; Smyrniotis, P. Z.; Ginsburg, A.; Stadtman, E. R. Metal Ion Requirement by Glutamine Synthetase of Escherichia Coli in Catalysis of γ-Glutamyl Transfer. Archives of Biochemistry and Biophysics 1975, 166 (1), 102–124.
- Moreira, C., Ramos, M. J., & Fernandes, P. A. (2017). Clarifying the Catalytic Mechanism of Human Glutamine Synthetase: A QM/MM Study. The Journal of Physical Chemistry B,121(26), 6313-6320. doi:10.1021/acs.jpcb.7b02543
- Berlicki, Ł. Inhibitors of Glutamine Synthetase and Their Potential Application in Medicine. Mini Rev Med Chem 2008, 8 (9), 869–878.
- Wired Chemist. (n.d.). Retrieved April 17, 2018, from http://www.wiredchemist.com/chemistr...metallic-radii
- Gill, H. S.; Eisenberg, D. The Crystal Structure of Phosphinothricin in the Active Site of Glutamine Synthetase Illuminates the Mechanism of Enzymatic Inhibition †. Biochemistry 2001, 40 (7), 1903–1912.
- Krajewski, W. W.; Collins, R.; Holmberg-Schiavone, L.; Jones, T. A.; Karlberg, T.; Mowbray, S. L. Crystal Structures of Mammalian Glutamine Synthetases Illustrate Substrate-Induced Conformational Changes and Provide Opportunities for Drug and Herbicide Design. Journal of Molecular Biology 2008, 375 (1), 217–228.
- Liaw, S. H.; Eisenberg, D. Structural Model for the Reaction Mechanism of Glutamine Synthetase, Based on Five Crystal Structures of Enzyme-Substrate Complexes. Biochemistry 1994, 33 (3), 675–681.
- Yang, X., Li, G., Yang, X., Jia, Z., & Luo, N. (n.d.). Determination of Manganese in Environmental Samples by UV-Vis after Cloud Point Extraction. INORGANIC CHEMISTRY,52(3), 1515-1524. doi:10.1021/ic302268h
Contributed by
This work was originally written by Kirstin Favazzo, Spring 2018: Kirstin is currently (as of 2018) a junior chemistry major at Saint Mary's College in Notre Dame, IN. This work was originally edited by Dr. Kathryn Haas (Assistant Professor), Madison Sendzik (Teaching and Research Assistant), and Dr. Dorothy Feigl (Professor) at Saint Mary's College.