
9.1: Biogeochemical Evolution
- Page ID
- 291428

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Present evidence suggests that blue-green algae, and possibly other primitive microbial forms of life, were flourishing 3 billion years ago. This brackets the origin of life to within one billion years; prior to 4 billion years ago, surface temperatures were probably above the melting point of iron, and there was no atmosphere nor hydrosphere.
By about 3.8 billion years ago, or one billion years after the earth was formed, cooling had occurred to the point where rain was possible, and primitive warm, shallow oceans had formed. The atmosphere was anoxic and highly reducing, containing mainly CO2, N2, CO, H2O, H2S, traces of H2, NH3, CH4, and less than 1% of the present amount of O2, probably originating from the photolysis of water vapor. This oxygen would have been taken up quite rapidly by the many abundant oxidizable substances such as Fe(II), H2S, and the like.
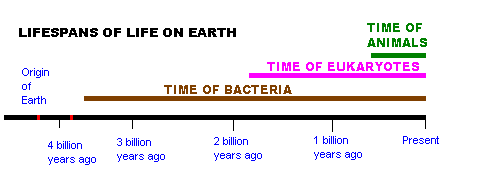
Timeline for development of the major life forms.
Evidence for early life
The fossil record that preserves the structural elements of organisms in sedimentary deposits has for some time provided a reasonably clear picture of the evolution of life during the past 750,000 years. In more recent years, this record has been considerably extended, as improved techniques have made it possible to study the impressions made by single-celled microorganisms embedded in rock formations.
The main difficulty in studying fossil microorganisms extending back beyond a billion years is in establishing that the relatively simple structural forms one observes are truly biogenic. There are three major kinds of evidence for this.
- Many of the most primitive life forms are still thriving, and these provide useful models with which some of the fossil forms can be compared.
- Carbon isotope ratios provide a second independent line of evidence for early life, or at least that of photosynthetic origin. In photosynthesis, C12O2 is taken up slightly more readily than is the heavier (and rarer) isotope C13O2; thus all but the very earliest life forms have left an isotopic fossil record even though the structural fossil may no longer be identifiable.
- A third evidence for early life is any indication of the presence of free oxygen in the local environment. Easily oxidizable species such as Fe(II) were very widely distributed on the primitive earth, and could not remain in contact with oxygen for very long without being oxidized. The oldest known formations of oxidized iron pyrite and of uranite are in sediments that were laid down between 2.0 and 2.3 billion years ago.
If all three of these lines of evidence are present in samples that can be shown to be contemporaneous with the sediments in which they are found, then the argument for life is incontrovertible. One of the most famous of these sites was discovered near Thunder Bay, Ontario in the early 1950’s. The Gunflint Formation consists of an exposed layer of chert (largely silica) from which the overlying shale of the Canadian Shield had been removed. Microscopic examination of thin sections of this rock revealed a variety of microbial cell forms, including some resembling present freshwater blue-green algae. Also present in the Gunflint Deposits are the oldest known examples of metazoa, or organisms which display a clear differentiation into two or more types of cell. These deposits have been dated at 1.9-2.0 billion years.

These filaments are believed to be the fossilized imprints of blue-green algae, one of the earliest life forms. They occur in the Bitter Springs Formation in Australia and are about 850 million years old.
The evidence from very old paleomicrobiotic deposits is less clear. Western Australia has yielded fossil forms that are apparently 2.8 billion years old, and other deposits in the same region contain structures resembling living blue-green algae. Other forms, heavily modified by chemical infiltration, bear some resemblence to a present iron bacterium, and are found in sediments laid down 3.5 billion years ago, but evidence that these fossils are contemporaneous with the sediments in which they are found is not convincing.
The oldest evidence of early life is the observed depletion of C13 in 3.8-billion year old rocks found in southwestern Greenland.
Origin of life: the organic environment
Under the conditions that prevailed at this time, most organic molecules would be thermodynamically stable, and there is every indication that a rich variety of complex molecules would be present. The most direct evidence of this comes from laboratory experiments that attempt to simulate the conditions of the primitive environment of this period, the first and most famous of these being the one carried out by Stanley Miller in 1953.
Schematic of the Miller experiment. A mixture of the various reduced gases believed to be compose the early atmosphere circulate through an apparatus in which spark discharges (intended to simulate lightning) create a complex mixture of organic compounds. S.L. Miller: A production of amino acids under possible primitive earth conditions. 1953: Science (117) 528-529. U. Texas provided the above diagram, and contains a very good description of of primitive life with many pictures of fossil organisms.
Since that time, other experiments of a similar nature have demonstrated the production of a wide variety of compounds under prebiotic conditions, including nearly all of the monomeric components of the macromolecules present in living organisms. In addition, small macromolecules, including peptides and sugars, as well as structural entities such as lipid-based micelles, have been prepared in this way.
The discovery in 1989 of a number of amino acids in the iridum-rich clay layer at the Cretaceous-Tertiary boundary suggests that bio-precursor molecules can be formed or deposited during a meteoric impact. Although this particular event occurred only 65 million years ago (and is presumed to be responsible for the extinction of the dinosaurs), the Earth has always been subject to meteoric impacts, and it is conceivable that these have played a role in the origin of life.
The presence of clays, whose surfaces are both asymmetric and chemically active, could have favored the formation of species of a particular chirality; a number of experiments have shown that clay surfaces can selectively adsorb amino acids which then form small peptides. It has been suggested that the highly active and ordered surfaces of clays not only played a crucial role in the formation of life, but might have actually served as parts of the first primitive self-replicating life forms, which only later evolved into organic species.
The first organisms
Since no laboratory experiment has yet succeeded in producing a self-replicating species that can be considered living, the mechanism by which this came about in nature must remain speculative. Infectious viruses have been made in the laboratory by simply mixing a variety of nucleotide precursors with a template nucleic acid and a replicase enzyme; the key to the creation of life is how to do the same thing without the template and the enzyme.
Smaller polynucleotides may have formed adventitiously, possibly on the active surface of an inorganic solid. These could form complementary base-paired polymers, which might then serve as the templates for larger molecules. Non-enzymatic template-directed synthesis of nucleotides has been demonstrated in the laboratory, but the resulting polymers have linkages that are not present in natural nucleotides.
It has been suggested that these linkages could have been selectively hydrolyzed by a long period of cycling between warm, cool, wet, and dry environmental conditions. The earth at that time was rotating more rapidly than it is now; cycles of hydration-dehydration and of heating-cooling would have been more frequent and more extreme.
The first organisms would of necessity have been heterotrophs— that is, they derived their metabolic energy from organic compounds in the environment. Their capacity to synthesize molecules was probably very limited, and they would have had to absorb many key substances from their surroundings in order to maintain their metabolic activity. Among the most primitive organisms of this kind are the archaeons, which are believed to be predecessors of both bacteria and eucaryotes. DNA sequencing of one such organism, a methane-producer that lives in ocean-bottom sediments at 200 atm and 48-94°C, reveals that only about a third of the genes resemble those of bacteria or eucaryotes.
It has been estimated that about 50 genes are required in order to define the minimal biochemical and structural machinery that a hypothetical simplest possible cell would have.
Development of photosynthetic organisms
The earliest organisms derived their metabolic energy from the organic substances present in their environment; once they began to reproduce, this nutrient source began to become depleted.
Some species had probably by this time developed the ability to reduce carbon dioxide to methane; the hydrogen source could at first have been H2 itself (at that time much more abundant in the atmosphere), and later, various organic metabolites from other species could have served.
Before the food supply neared exhaustion, some of these organisms must have developed at least a rudimentary means of absorbing sunlight and using this energy to synthesize metabolites. The source of hydrogen for the reduction of CO2 was at first small organic molecules; later photosynthetic organisms were able to break this dependence on organic nutrients and obtain the hydrogen from H2S.
These bacterial forms were likely the dominant form of life for several hundred million years. Eventually, due perhaps to the failing supply of H2S, plants capable of mediating the photochemical extraction of hydrogen from water developed. This represented a large step in biochemical complexity; it takes 10 times as much energy to abstract hydrogen from water than from hydrogen sulfide, but the supply is virtually limitless.
It appears that photosynthesis evolved in a kind of organism whose present-day descendents are known as cyanobacteria.
Procaryotes and eucaryotes
The five “kingdoms” into which living organisms are classified are Monera, Protista (protozoans, algae), Fungi, Plantae, and Animalia. The genetic (and thus, evolutionary) relations between these and the subcategories within them are depicted below.
Superimposed on this, however, is an even more fundamental division between the procaryotes and eucaryotes.
Procaryotes
In this group are primitive organisms whose single cells contain no nucleus; the gene-bearing structure is a single long DNA chain that is folded irregularly throughout the cell. Procaryotic cells usually reproduce by budding or division; where sexual reproduction does occur, there is a net transfer of some genetic material from one cell to another, but there is never an equal contribution from both parents.
In spite of their primitive nature, procaryotes constitute the majority of organisms in the biosphere. The division between bacteria and archaea within the procaryotic group is a fairly recent one. Archaea are now believed to be the most primitive of all organisms, and include the so-called extremophiles that occupy environmental niches in which life was at one time thought to be impossible; they have been found in sedimentary rocks, hot springs, and highly saline environments.
Eucaryotes.
All other organisms— seaweeds (algae), protozoa, molds, fungi, animals and plants, are composed of eucaryotic cells. These all have a membrane-bound nucleus, and with a few exceptions they all reproduce by mitosis, in which the chromosomes split longitudinally and move toward opposite poles. Other organelles unique to eucaryotes are mitochondria, ribosomes, and structural elements such as microtubules.
Oxygen and biogeochemical evolution
Oxygen is poisonous to all forms of life in the absence of enzymes that can reduce the highly reactive byproducts of oxidation and oxidative metabolism (peroxides, superoxides, etc.). All organic compounds are thermodynamically unstable in the presence of oxygen; carbon-carbon double bonds in lipids are subject to rapid attack. Prebiotic chemical evolution leading to the development of biopolymers was possible only under the reducing, anoxic conditions of the primitive atmosphere.
The rise of atmospheric oxygen. Once organisms existed that could use water as a hydrogen source for the reduction of carbon dioxide, O2began to be introduced into the atmosphere. The widespread occurrence of ferrous compounds in surface rocks and sediments provided a sink for this oxygen that probably did not become saturated until about 2 billion years ago, when the atmospheric oxygen abundance first rose above about 1 percent.Illustration from All about entropy, the laws of thermdynamics, and order from disorder. There is lots of good stuff here!
As the oxygen concentration began to rise, organisms in contact with the atmosphere had to develop protective mechanisms in order to survive. One indication of such adaptation is the discovery of fossil microbes whose cell walls are unusually thick. A more useful kind of adaptation was the synthesis of compounds that would detoxify oxygen by reacting rapidly either with O2 itself or with peroxides and other active species derived from it. Isoprenoids (the precursors of steroids) and porphyrins are examples of two general classes of compounds that are found in nearly all organisms, and which may have originated in this way. Later, highly efficient oxygen mediating enzymes such as peroxidase and catalase developed. The widespread phenomenon of bioluminescence may be the result of a very early adaptation to oxygen. The compound luciferin is a highly efficient oxygen detoxifier, which also happens to be able to emit light under certain conditions. Bioluminescence probably developed as a by-product in early procaryotic organisms, but was gradually lost as more efficient detoxifying mechanisms became available.
Development of the eucaryotes
In spite of the deleterious effects of oxygen on cell biomolecules, O2 is nevertheless an excellent electron sink, capable of releasing large quantities of energy through the oxidation of glucose. This energy can be efficiently captured through oxidative phosphorylation, the key process in respiration.
A cell that utilizes oxygen must have a structural organization that isolates the oxygen-consuming respiratory centers from the other parts of the cell that would be poisoned by oxygen or its reaction products. Some procaryotic organisms have developed in this way; a number of cyanobacteria and other species are facultative anaerobes which can survive both in the presence and absence of oxygen.
It is in the eucaryotic cell, however, that this organization is fully elaborated; here, respiration occurs in membrane-bound organelles called mitochondria. With only a few exceptions, all eucaryotic organisms are obligate aerobes; they can rarely survive and can never reproduce in the absence of oxygen. Mitotic cell division depends on the contractile properties of the protein actomyosin, which only forms when oxygen is present.
The development of the eucaryotic cell about 1.4 billion years ago is regarded as the most significant event in the evolution of the earth and of the biosphere since the appearance of photosynthesis and the origin of life itself. How did it come about? The present belief, supported by an increasing amount of evidence, suggests that it began when one species of organism engulfed another. The ingested organism possessed biochemical machinery not present in the host, but which was retained in such a way that it conferred a selective evolutionary advantage on the host. Eventually the two organisms became able to reproduce as one, and so effectively became a single organism. This process is known as endosymbiosis
According to this view, mitochondria represent the remains of a primitive oxygen-tolerant organism that was incorporated into one that could produce the glucose fuel for the oxygen to burn. Chloroplasts were once free-living photosynthesizing procaryotes similar to present-day cyanobacteria. It is assumed that some of these began parasitising respiratory organisms, conferring upon them the ability to synthesize their carbohydrate food during daylight. The immense selective advantage of this arrangement is evident in the extent of the plant kingdom.
Consequences of the oxygen increase
It is interesting that an atmospheric oxygen concentration of about 1 percent, known as the Pasteur point is both the maximum that obligate anaerobes can tolerate, and the minimum required for oxidative phosphorylation.
Louis Pasteur discovered that some bacteria are anaerobic and unable to tolerate oxygen above 1% concentration.
As was mentioned previously, the oxygen produced by the first photosynthetic organisms was taken up by ferrous iron in sediments and surface minerals. The widespread deposits known as banded iron formations consist of alternating layers of Fe(III)-containing oxides (hematite and magnetite) that were laid down between 1 and 2 billion years ago; the layering may reflect changing climatic or other environmental conditions that brought about a cycling of the organism population.
During the buildup of oxygen, an equivalent amount of carbon had to be deposited in sediments in order to avoid the thermodynamically spontaneous back reaction which would consume the O2through oxidation of the organic matter. Thus the present levels of atmospheric oxygen are due to a time lag in the geochemical cycling of photosynthetic products.
As the oxygen concentration increased, evolution seems to have speeded up; this may reflect both the increased metabolic efficiency and the greater biochemical complexity of the eucaryotic cell. The oldest metazoan (multiple-celled) fossils are coelenterates that appeared about 700 million years ago. Modern representatives of this group such as marine worms and jellyfish can tolerate oxygen concentrations as low as 7%, thus placing a lower boundary on the atmospheric oxygen content of that era. The oldest fossil organisms believed to have possessed gills, which function only above 10% oxygen concentration, appeared somewhat later.Carbon dioxide decreased as oxygen increased, as indicated by the prevalence of dolomite over limestone in early marine sediments.