
11.8: Flavin-Dependent Monooxygenase Reactions - Hydroxylation, Epoxidation, and the Baeyer-Villiger Oxidation
- Page ID
- 242310

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
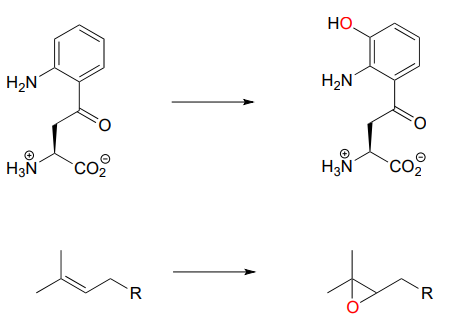
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Up to now, the redox reaction examples we have seen have all been either hydrogenation/dehydrogenation transformations or interconversions between free thiols and disulfides. However, there are many important redox reactions in biological chemistry which do not fall under either of these descriptions. Oxygenase enzymes catalyze the insertion of one or two oxygen atoms from molecular oxygen (\(O_2\)) into an organic substrate molecule. Enzymes which insert a single oxygen atom are called monooxygenases. Below are two examples of biochemical transformations catalyzed by monooxygenase enzymes: one is a hydroxylation, the other is an epoxidation (an epoxide functional group is composed of a three-membered carbon-carbon-oxygen ring - epoxides are somewhat rare in biological organic chemistry but are very common and useful intermediates in laboratory organic synthesis).
Dioxygenase enzymes insert both oxygen atoms from \(O_2\) into the substrate, and usually involve cleavage of an aromatic ring. Below is an example of a dioxygenase reaction, catalyzed by catechol dioxygenase:

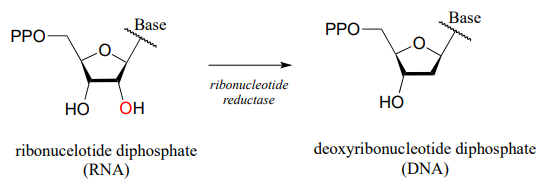
In the reduction direction, reductases remove oxygen atoms, or sometimes other electronegative heteratoms such as nitrogen or halides. For example, DNA deoxyribonucleosides are converted from their corresponding RNA ribonucleosides by the action of reductase enzymes:
Many oxygenase and reductase reactions involve the participation of enzyme-bound transition metals - such as iron or copper - and the mechanistic details of these reactions are outside the scope of our discussion. A variety of biochemical monooxygenase reactions, however, involve flavin as a redox cofactor, and we do have sufficient background knowledge at this point to understand these mechanisms. In flavin-dependent monooxygenase reactions, the key intermediate species is flavin hydroperoxide.
The term 'peroxide' refers to a functional group characterized by an oxygen-oxygen single bond. The simplest peroxide is hydrogen peroxide (\(HOOH\)) about which we will have more to say below. In flavin hydroperoxide, the peroxide group is linked to one of the carbons of the reactive triple-ring system of the coenzyme. A possible mechanism for the formation of flavin peroxide from \(FADH_2\) and molecular oxygen is shown below.
Silverman, R.B. The Organic Chemistry of Enzyme-Catalyzed Reactions, p. 121-122, Scheme 3.33. 2000, Academic Press, San Diego.
Mechanism for the formation of flavin hydroperoxide:
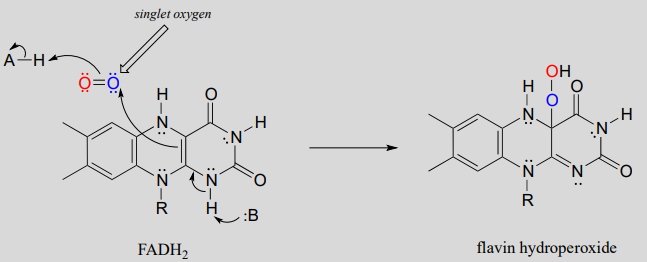
(Note: Implicit in this mechanism is that the molecular oxygen first undergoes spin inversion from the triplet state to the higher energy 'singlet' state. You may recall from your general chemistry course that molecular oxygen exists in two states: 'singlet' oxygen has a double bond and no unpaired electrons, while 'triplet' oxygen has a single \(O-O\) bond and two unpaired electrons - a kind of 'double radical'. Molecular orbital theory - and experimental evidence - show that the triplet state is lower in energy.
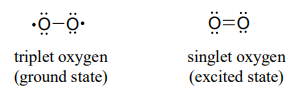
The mechanism shown above is one proposed mechanism, another proposal involves triplet oxygen reacting with flavin in a series of radical-intermediate, single-electron steps.)
Flavin hydroperoxide can be thought of as an activated form of molecular oxygen. Peroxides in general are potent oxidizing agents, because the oxygen-oxygen single bond is quite weak: only 138 kJ/mole, compared to 339 kJ/mol for a carbon-carbon bond, and 351 kJ/mol for a carbon-oxygen bond. When the 'outer' oxygen of flavin hydroperoxide (red in our figure above) comes into close proximity to the p-bonded electrons of an alkene or aromatic group, the \(O-O\) bond will break, leaving an empty orbital on the outer oxygen to be filled by the p electrons - thus, a new carbon-oxygen bond is formed. This is what is happening in step 1 of a reaction in the tryptophan degradation pathway catalyzed by kynurenine 3-monooxygenase. Step 2 completes what is, mechanistically speaking, an electrophilic aromatic substitution reaction (section 14.4) with an peroxide oxygen electrophile.
Mechanism for the flavin hydroperoxide-dependent hydroxylation of kynurenine:
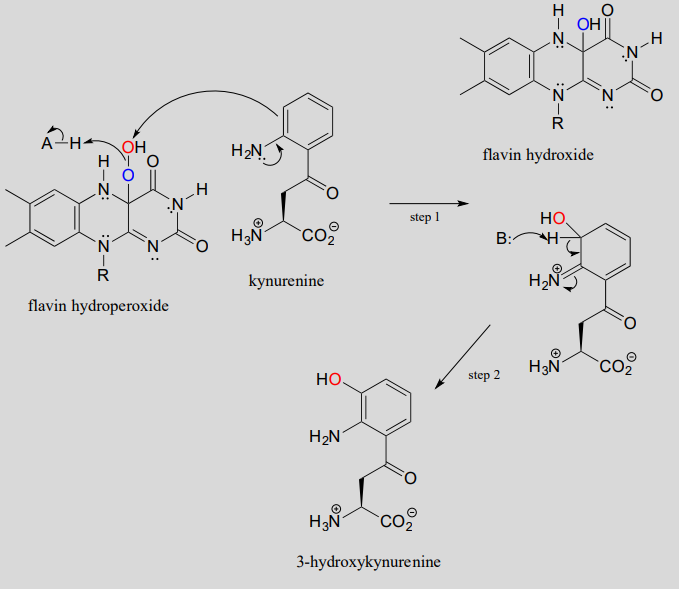
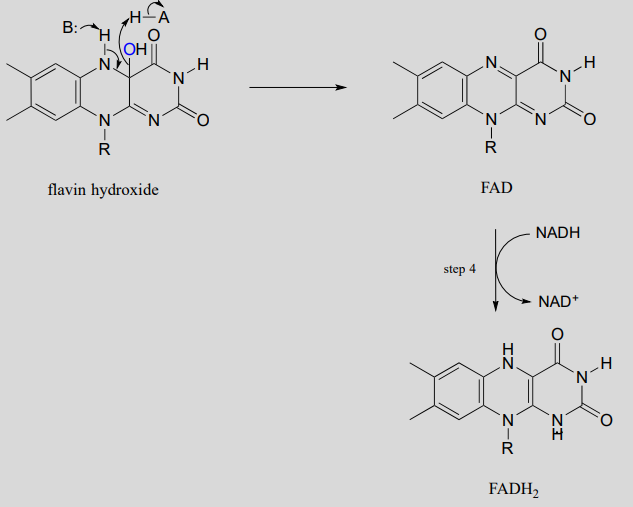
Elimination of water from the hydroxyflavin intermediate then leads to formation of \(FAD\) (step 3), which is subsequently reduced back to \(FADH_2\) by \(NADH\) (step 4).
The \(N\)-hydroxylation reaction below, which is part of the of the biosynthetic pathway of an iron-binding molecule in the pathogenic bacterium Pseudomonas aeruginosa, is mechanistically similar to the \(C\)-hydroxylation reaction we just saw, except that the nucleophile is an amine nitrogen. Note that \(FADH_2\) is shown in brackets below the reaction arrow, indicating that reduced flavin participates in the reaction but is not used up - rather it is regenerated in the active site at the end of the reaction cycle.
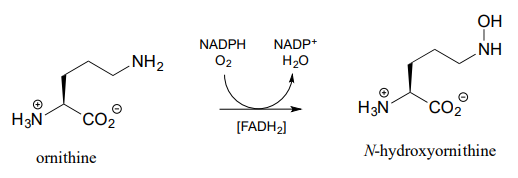
Draw arrows for the \(N-O\) bond-forming step in the ornithine hydroxylation reaction above.
Epoxides, characterized by a three-membered ring composed of two carbons and one oxygen, are a very common and useful functional group employed in synthetic organic chemistry. Although rare, there are some interesting epoxide-forming reactions in biochemical pathways, catalyzed by flavin-dependent monooxygenase enzymes.
In a key step in the biosynthesis of cholesterol and other steroid compounds, an alkene is converted to an epoxide in a precursor molecule called squalene. Flavin hydroperoxide also serves as the direct oxidizing agent in this step:
Mechanism for the flavin-hydroperoxide-dependent epoxidation of squalene:

Oxidosqualene goes on to cyclize to lanosterol in a complex and fascinating electrophilic reaction which we discussed in section 14.5.
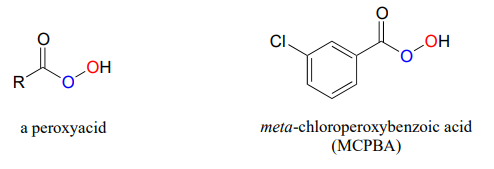
Epoxidation reactions have a parallel in the synthetic organic laboratory, and in fact are very important tools in organic synthesis. In laboratory epoxidations, peroxyacids are the counterpart to flavin hydroperoxide in biochemical epoxidations. meta-chloroperoxybenxoic acid (MCPBA) is a commonly used peroxyacid.
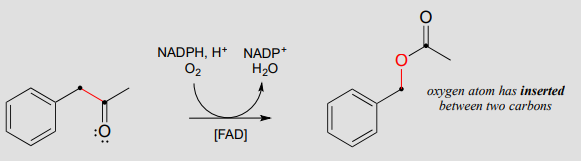
The Baeyer-Villiger oxidation, in which a ketone is converted to an ester through treatment with a peroxide reagent, is an extremely useful laboratory organic synthesis reaction discovered in the late 19th century. Recently, many biochemical examples of Baeyer-Villiger oxidations have been discovered: the reaction below, for example, is catalyzed by a monooxygenase in a thermophilic bacterium:
(Proc. Natl. Acad. Sci. U.S.A. 2004, 101, 13157)
A Baeyer-Villiger oxidation:
Mechanism:
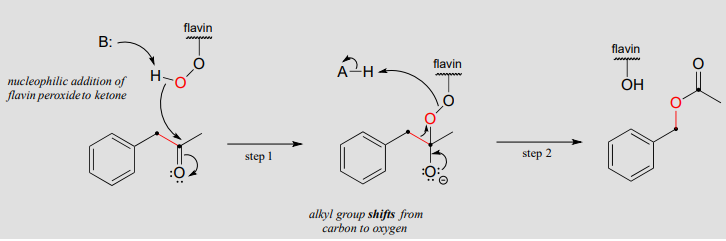
The Baeyer-Villiger mechanism is differs significantly from the hydroxylation reactions we saw earlier, although flavin hydroperoxide (abbreviated in the above figure) still plays a key role. Here, the peroxide oxygen is a nucleophile, rather than an electrophile, attacking the ketone carbonyl in step 1. Step 2 is a rearrangement, similar in many ways to the hydride and alkyl shifts we learned about in section 14.5. The electrons in the red bond in the figure shift over one atom: from the carbonyl carbon to the outer peroxide oxygen. The end result is that an oxygen atom, from \(O_2\) via flavin hydroperoxide, has been inserted between the carbonyl carbon and a neighboring methylene (\(CH_2\)) carbon, forming an ester.
Note that in the reaction mechanism above, the ketone substrate is asymmetric: on one side of the carbonyl there is a benzyl group (\(CH_2\)-phenyl), and on the other side a methyl group. Note also that it is the benzyl group, not the methyl, that shifts in step 2 of the mechanism. For reasons that are not yet well understood, in Baeyer-Villiger reactions the alkyl group with higher carbocation stability has a higher migratory aptitude: in other words, it has a lower energy barrier for the shifting step.
Draw the product of a hypothetical Baeyer-Villiger reaction involving the same substrate as the above figure, in which the methyl rather than the benzyl group shifts.
Draw the likely major product of a hypothetical Baeyer Villiger reaction starting with 2-methylcyclopentanone as the substrate. Take into account the idea of migratory aptitude.
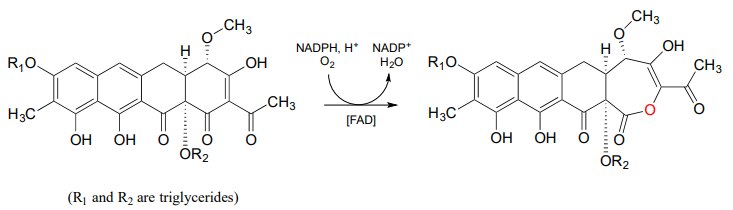
Below is another example of a Baeyer-Villiger reaction in which a cyclic ketone is oxidized to a lactone (cyclic ester). Notice that oxygen insertion expands the ring from 6 to 7 atoms. This is the third-to-last step in the biosynthesis of the anti-cancer agent mithromycin in some bacterial species (ACS Chem. Biol. 2013, 8, 2466).
Yet another variety of flavin-dependent monooxygenase, which bears some mechanistic similarity to the Baeyer-Villiger oxidation, is the decarboxylative reaction below from biosynthesis of the plant hormone auxin: (J. Biol. Chem. 2013, 288, 1448)
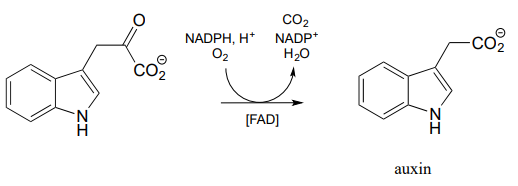
Propose a mechanism for the above reaction, starting with flavin hydroperoxide.