12.11: Carbohydrates Summary
- Page ID
- 338378

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The last class of macromolecules we will consider structurally here is the carbohydrates. Built of sugars or modified sugars, carbohydrates have several important functions, including structural integrity, cellular identification, and energy storage.
Monosaccharides
Simple sugars, also known as monosaccharides, can generally be written in the form \(C_x(H_2O)_x\). It is for this reason they are referred to as carbo-hydrates. By convention, the letters ‘ose’ at the end of a biochemical name flags a molecule as a sugar. Thus, there are glucose, galactose, sucrose, and many other ‘-oses’. Other descriptive nomenclature involves use of a prefix that tells how many carbons the sugar contains. For example, glucose, which contains six carbons, is described as a hexose. The following list shows the prefixes for numbers of carbons in a sugar:
- Tri- = 3
- Tetr- = 4
- Pent- = 5
- Hex- =6
- Hept- = 7
- Oct- = 8
Other prefixes identify whether the sugar contains an aldehyde group (aldo-) or a ketone (keto) group. Prefixes may be combined. Glucose, which contains an aldehyde group, can be described as an aldo-hexose. The list that follows gives some common sugars and some descriptors.
- Ribose = aldo-pentose
- Glucose = aldo-hexose
- Galactose = aldo-hexose
- Mannose = aldo-hexose
- Glyceraldehyde = aldo-triose
- Erythrose – aldo-tetrose
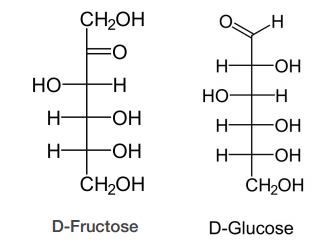
- Fructose = keto-hexose
- Ribulose = keto-pentose
- Sedoheptulose = keto-heptose
- Dihydroxyacetone = keto-triose
Stereoisomer Nomenclature
Sugars of a given category (hexoses, for example) differ from each other in the stereoisomeric configuration of their carbons. Two sugars having the same number of carbons (hexoses, for example) and the same chemical form (aldoses, for example), but differing in the stereoisomeric configuration of their carbons are called diastereomers. Biochemists use D and L nomenclature to describe sugars, as explained below.
D-sugars predominate in nature, though L-forms of some sugars, such as fucose, do exist. The D and L designation is a bit more complicated than it would appear on the surface. To determine if a sugar is a D-sugar or an L-sugar, one simply examines the configuration of the highest numbered asymmetric carbon. If the hydroxyl is written to the right, it is a D-sugar. If the hydroxyl is on the left, it is an L-sugar. That part is simple. The confusion about D and L arises because L sugars of a given name (glucose, for example) are mirror images of D sugars of the same name. The figure on the previous page shows the structure of D- and L- glucose. Notice that D-glucose is not converted into L-glucose simply by .ipping the configuration of the fifth carbon in the molecule. There is another name for sugars that are mirror images of each other. They are called enantiomers. Thus, L-glucose and D-glucose are enantiomers, but D-Erythrose and D-Threose are diastereomers.


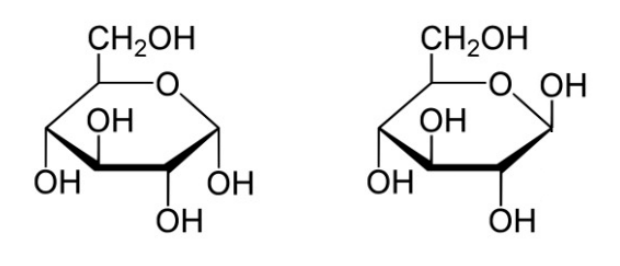
Sugars of 5-7 carbons can fairly easily form ring structures (called Haworth structures). For aldoses like glucose, this involves formation of a hemi-acetal. For ketoses like fructose, it involves formation of a hemi-ketal. The bottom line for both is that the oxygen that was part of the aldehyde or the ketone group is the one that becomes a part of the ring. More important than the oxygen, though, is the fact that the carbon attached to it (carbon #1 in aldoses or #2 in ketoses) becomes asymmetric as a byproduct of the cyclization. This new asymmetric carbon is called the anomeric carbon and it has two possible configurations, called alpha and beta.
A solution of glucose will contain a mixture of alpha and beta forms. Whether the alpha or the beta arises upon cyclization is partly determined by geometry and partly random. Thus, one can find a bias for one form, but usually not that form exclusively. A given molecule of sugar will flip between alpha and beta over time. A requirement for this is that the hydroxyl on the anomeric carbon is unaltered, thus facilitating flipping back to the straight chain form followed by recyclization. If the hydroxyl becomes chemically altered in any way (for example, replacement of its hydrogen by a methyl group), a glycoside is formed. Glycosides are locked in the same alpha or beta configuration they were in when the modification was made. Glycosides are commonly found in nature. Sucrose, for example , is a di-glycoside – both the glucose and the fructose have had their anomeric hydroxyls altered by being joined together.
The last considerations for sugars relative to their structure are their chemical reactivity and modification. The aldehyde group of aldoses is susceptible to oxidation, whereas ketoses are less so. Sugars that are readily oxidized are called ‘ reducing sugars ’ because their oxidation causes other reacting molecules to be reduced. Reducing sugars can easily be identified in a chemical test. Chemical modification of sugars occurs readily in cells. As we will see, phosphorylation of sugars occurs routinely during metabolism. Oxidation of sugars to create carboxyl groups also can occur. Reduction of aldehyde/ketone groups of sugars creates what are called sugar alcohols, and other modifications, such as addition of sulfates and amines also readily occur.

Disaccharides
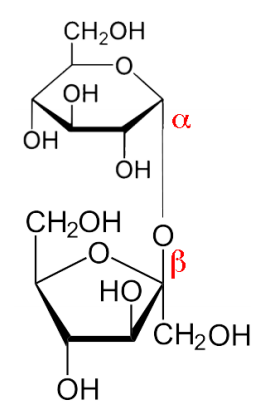
Sugars are readily joined together (and broken apart) in cells. Sucrose (Figure \(\PageIndex{7}\)), which is common table sugar, is made by joining the anomeric hydroxyl of alpha-D-glucose to the anomeric hydroxyl of beta-D-fructose. Not all disaccharides join the anomeric hydroxyls of both sugars. For example, lactose (milk sugar) is made by linking the anomeric hydroxyl of galactose in the beta configuration to the hydroxyl of carbon #4 of glucose.
Polysaccharides
Polysaccharides, as their name implies, are made by joining together many sugars. The functions for polysaccharides are varied. They include energy storage, structural strength, and lubrication. Polysaccharides involved in energy storage include the plant polysaccharides, amylose and amylopectin. The polysaccharide involved in energy storage in animals is called glycogen and it is mostly found in the muscles and liver.
Amylose/Amylopectin
Amylose is the simplest of the polysaccharides, being comprised solely of glucose units joined in an alpha 1-4 linkage. Amylose is broken down by the enzyme alpha-amylase, found in saliva. Amylopectin is related to amylose in being composed only of glucose, but it differs in how the glucose units are joined together. Alpha 1-4 linkages predominate, but every 30-50 residues, a ‘branch’ arises from an alpha 1-6 linkage. Branches make the structure of amylopectin more complex than that of amylose.
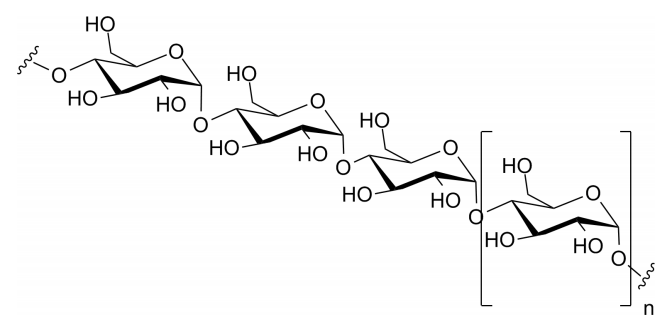
Glycogen
Glycogen is a polysaccharide that is physically related to amylopectin in being built only of glucose and in having a mix of alpha 1-4 and alpha 1-6 bonds. Glycogen, however, has many more alpha 1-6 branches than amylopectin, with such bonds occurring about every 10 residues. One might wonder why such branching occurs more abundantly in animals than in plants. A plausible explanation is based on the method by which these molecules are broken down. The breakdown of these polysaccharides is catalyzed by enzymes, known as phosphorylases, that clip glucose residues from the ends of glycogen chains and attach a phosphate to them in the process, producing glucose-1-phosphate. More highly branched polysaccharides have more ends to clip, and this translates to more glucose-1-phosphates that can be removed simultaneously by numerous phosphorylases. Since glucose is used for energy by muscles, glucose concentrations can be increased faster the more branched the glycogen is. Plants, which are immobile do not have needs for such immediate release of glucose and thus have less need for highly branched polysaccharides.
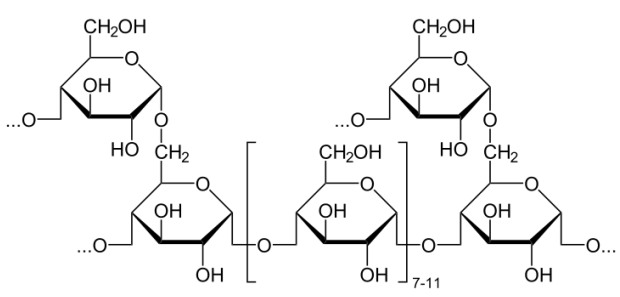
Cellulose
Another important polysaccharide containing only glucose is cellulose. It is a polymer of glucose used to give plant cell walls structural integrity and has the individual units joined solely in a beta 1-4 configuration. That simple structural change makes a radical diff erence in its digestibility. Humans are unable to break down cellulose and it passes through the digestive system as roughage. Ruminant animals, such as cattle, however have bacteria in their rumens that contain the enzyme cellulase. It breaks the beta 1-4 links of the glucoses in cellulose to release the sugars for energy.
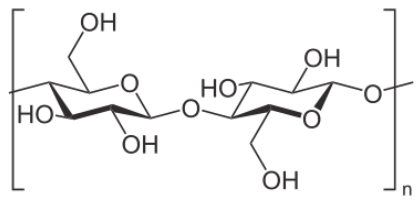
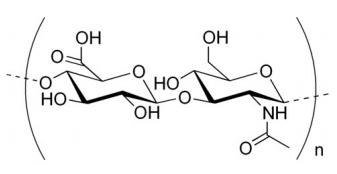
Another polysaccharide used for structural integrity is known as chitin. Chitin makes up the exoskeleton of insects and is a polymer of a modified form of glucose known as N-acetyl-glucosamine.
Glycosaminoglycans
Yet another category of polysaccharides are the glycosaminoglycans (also called mucopolysaccharides ), some examples of which include keratan sulfate, heparin, hyaluronic acid (right), and chondroitin sulfate. The polysaccharide compounds are linked to proteins, but differ from glycoproteins in having a much larger contingent of sugar residues and, further, the sugars are considerably more chemically modified. Each of them contains a repeating unit of a disaccharide that contains at least one negatively charged residue. The result is a polyanionic substance that, in its interactions with water, makes for a “slimy" feel. Glycosaminoglycans are found in snot, and in synovial fluid, which lubricates joints. Heparin is a glycosaminoglycan that helps to prevent blood from clotting.