
5.4.1: Optional deep dive in to membrane transport
- Page ID
- 354313

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Source: BiochemFFA_3_2.pdf. The entire textbook is available for free from the authors at http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Active transport
All of the transport mechanisms described so far are driven solely by a concentration gradient - moving from higher concentrations in the direction of lower concentrations. These movements can occur in either direction and, as noted, result in equal concentrations on either side of the bilayer, if allowed to go to completion. Many times, however, cells must move materials against a concentration gradient and when this occurs, another source of energy is required. This process is known as active transport.
A good definition of active transport is that during active transport, at least one molecule is being moved against a concentration gradient. A common, but not exclusive, energy source is ATP (see Na+/K+ ATPase), but other energy sources are also employed. For example, the sodium-glucose transporter uses a sodium gradient as a force for actively transporting glucose into a cell. Thus, it is important to know that not all active transport uses ATP energy.
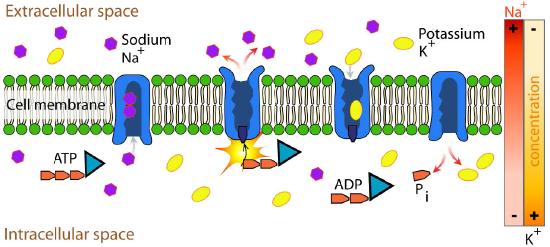
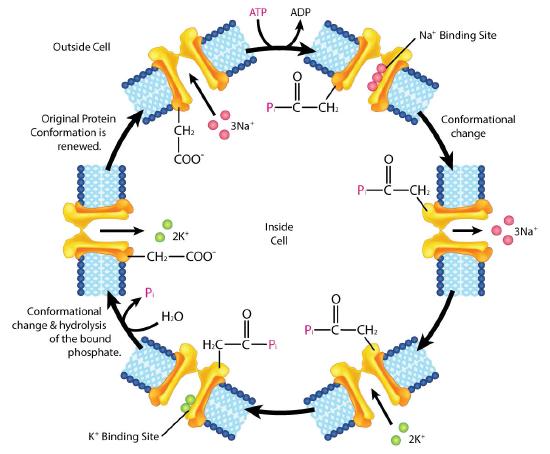
Na+/K+ ATPase
An important integral membrane transport protein is the Na+/K+ ATPase antiport (Figures 3.37 and 3.38), which moves three sodium ions out of the cell and two potassium ions into the cell with each cycle of action. In each case, the movement of ions is against the concentration gradient. Since three positive charges are moved out for each two positive charges moved in, the system is electrogenic.
The protein uses the energy of ATP to create ion gradients that are important both in maintaining cellular osmotic pressure and (in nerve cells) for creating the sodium and potassium gradients necessary for signal transmission. Failure of the system to function results in swelling of the cell due to movement of water into the cell through osmotic pressure. The transporter expends about one fifth of the ATP energy of animal cells. The cycle of action occurs as follows:
- Pump binds ATP followed by binding of 3 Na+ ions from cytoplasm of cell
- ATP hydrolysis results in phosphorylation of aspartate residue of pump. ADP is released
- Phosphorylated pump undergoes conformational change to expose Na+ ions to exterior of cell. Na+ ions are released.
- Pump binds 2 extracellular K+ ions.
- Pump dephosphorylates causing it to expose K+ ions to cytoplasm as pump returns to original shape.
- Pump binds 3 Na+ ions, binds ATP and releases 2 K+ ions to restart process
The Na+/K+ ATPase is classified as a P-type ATPase. This category of pump is notable for having a phosphorylated aspartate intermediate and is present across the biological kingdoms - bacteria, archaeans, and eukaryotes.
ATPase types
ATPases have roles in either the synthesis or hydrolysis of ATP and come in several different forms.
- F-ATPases (F1FO-ATPases) are present in mitochondria, chloroplasts and bacterial plasma membranes and are the prime ATP synthesizers for these systems. Each uses a proton gradient as its energy source for ATP production. Complex V of the mitochondrion is an F-type ATPase.
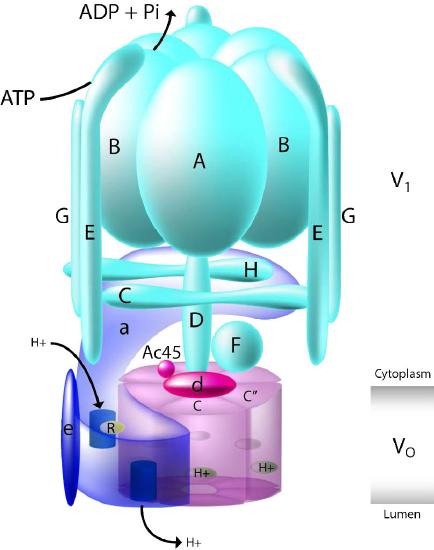
- V-ATPases (V1VO-ATPases) are mostly found in vacuoles of eukaryotes . They utilize energy from ATP hydrolysis to transport solutes and protons into vacuoles and lysosomes, thus lowering their pH values.
The V-type and F-type ATPases are very similar in structure. The V-type (Figure 3.39) uses ATP to pump protons into vacuoles and lysosomes, whereas F-types use proton gradients of the mitochondria and chloroplasts to make ATP.
- A-ATPases (A1AO-ATPases) are found in archaeans and are similar to F-ATPases in function.
- P-ATPases (E1E2-ATPases) are in bacteria, fungi and in eukaryotic plasma membranes and organelles. They transport a diversity of ions across membranes. Each has a common mechanism of action which include autophosphorylation of a conserved aspartic acid side chain within it. Examples of P-type ATPases include the Na+/K+ ATPase and the calcium pump.
- E-ATPases are enzymes found on the cell surface. They hydrolyze a range of extracellular nucleoside triphosphates, including ATP.
Nerve transmission
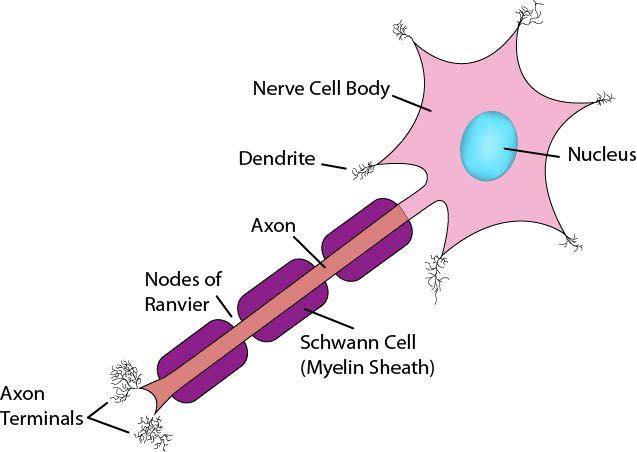
Now that you have seen how the Na+/K+ ATPase functions, it is appropriate to discuss how nerve cells use ion gradients created with it to generate and transmit nerve signals. Neurons are cells of the nervous system that use chemical and electrical signals to rapidly transmit information across the body (Figure 3.40). The sensory nerve system links receptors for vision, hearing, touch, taste, and smell to the brain for perception. Motor neurons run from the spinal cord to muscle cells. These neurons have a cell body and a very long, thin extension called an axon, that stretches from the cell body in the spinal cord all the way to the muscles they control. Nerve impulses travel down the axon to stimulate muscle contraction.
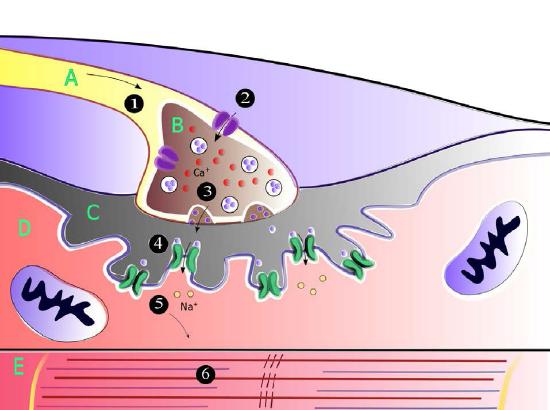
Signals travel through neurons, ultimately arriving at junctions with other nerve cells or target cells such as muscle cells. Note that neurons do not make physical contact with each other or with muscle cells. The tiny space between two neurons or between a neuron and a muscle cell is called the synaptic cleft. At the synaptic cleft, the neuron releases neurotransmitters that exit the nerve cell and travel across the junction to a recipient cell where a response is generated. That response may be creating another nerve signal, if the adjacent cell is a nerve cell or it may be a muscular contraction if the recipient is a muscle cell (Figure 3.41).
In considering information movement via nerve cells, then, we will discuss two steps - 1) creation and propagation of a signal in a nerve cell and 2) action of neurotransmitters exiting a nerve cell and transiting a synaptic junction.
Signal source
Creation of a nerve signal begins with a stimulus to the nerve cell. In the case of muscle contraction, the motor cortex of the brain sends signals to the appropriate motor neurons, stimulating them to generate a nerve impulse. How is such an impulse generated?
Resting potential
In the unstimulated state, all cells, including nerve cells, have a small voltage difference (called the resting potential) across the plasma membrane, arising from unequal pumping of ions across the membrane. The Na+/K+ ATPase, for example, pumps sodium ions out of the cell and potassium ions into cells. Since three sodium ions get pumped out for every two potassium ions pumped in, a charge and chemical gradient is created. It is the charge gradient that gives rise to the resting potential.
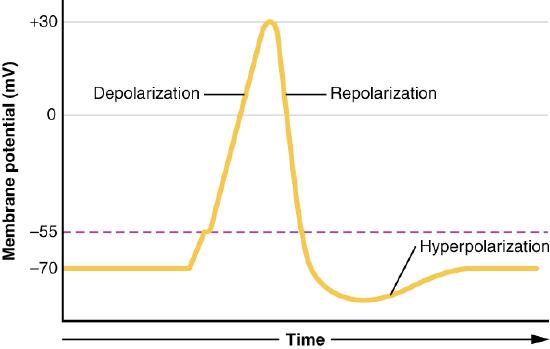
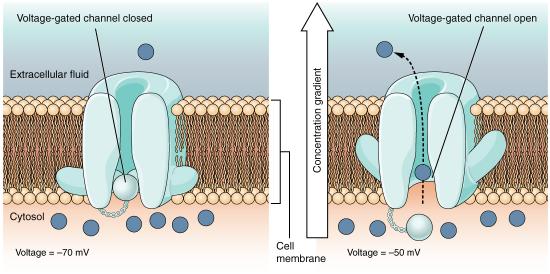
Altering the gradients of ions across membranes provide the driving force for nerve signals. This happens as a result of opening and closing of gated ion channels. Opening of gates to allow ions to pass through the membrane swiftly changes the ionic balance across the membrane resulting in a new voltage difference called the action potential. It is the action potential that is the impetus of nerve transmission.
Initiation of signal
The signal generated by a motor neuron begins with opening of sodium channels in the membrane of the nerve cell body causing a rapid influx of sodium ions into the nerve cell. This step, called depolarization (Figure 3.42), triggers an electrochemical signal - the action potential. Remember that the Na+/K+ ATPase has created a large sodium gradient, so sodium ions rush into the cell when sodium channels open. After the initial depolarization, potassium channel gates, responding to the depolarization, open, allowing potassium ions to rapidly diffuse out of the cell (remember K+ ions are more abundant inside of the cell). This phase is called the repolarization phase and during it, the sodium gates close.
The rapid exit of potassium ions causes the voltage difference to “overshoot” the resting potential and potassium gates close. This followed by the so-called refractory period, when the Na+/K+ ATPase begins its work to re-establish the original conditions by pumping sodium ions out and potassium ions into the nerve cell. Eventually, the system recovers and the resting potential is re-established. The initiating end of the nerve cell is then ready for another signal.
Propagation of action potential
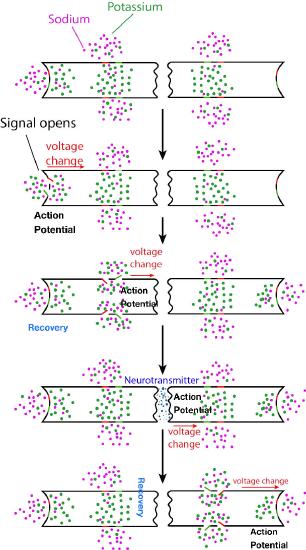
What we have described here is only the initiation of the nerve signal in one part of the nerve cell. For the signal to be received, the action potential must travel the entirety of the length of the nerve cell (the axon) and cause a chemical signal to be released into the synaptic cleft to get to its target. Propagating the nerve signal (action potential) in the original nerve cell is the function of all of the rest of the gated ion channels (Figure 3.43) positioned on the sides of the nerve cell. The sodium and potassium gates involved in propagation of the signal all act in response to voltage changes created by the electrochemical gradient moving down the nerve cell (Figure 3.44). Remember that opening of the initial gates at initiation of the signal created an influx of sodium ions and an efflux of potassium ions.
Moving signal
This chemical and electrical change that creates the action potential leaves the end of the nerve cell where it started and travels down the axon towards the other end of the nerve cell. Along the way, it encounters more sodium and potassium gated channels. In each case, these respond simply to the voltage change of the action potential and open and close, exactly in the same way the gates opened to start the signal. Thus, a rapid wave of increasing sodium ions and decreasing potassium ions moves along the nerve cell, propagated (and amplified) by gates opening and closing as the ions and charges move down the nerve cell. Eventually, the ionic tidal wave reaches the end of the nerve cell (axon terminal) facing the synaptic cleft.
Crossing the synaptic cleft
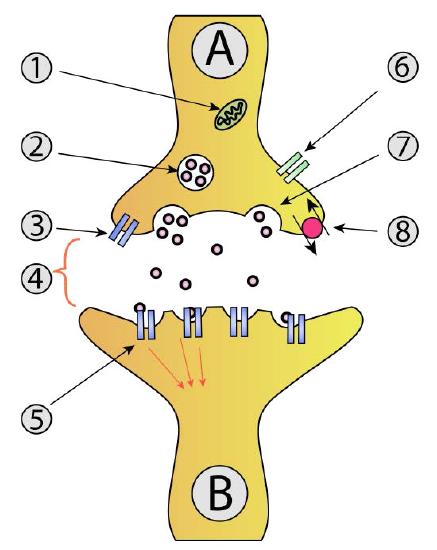
For the signal to be received by the intended target (postsynaptic cell) from the originating neuron (presynaptic neuron), it must cross the synaptic cleft and stimulate the neighboring cell (Figure 3.45). Communicating information across a synaptic cleft is the job of neurotransmitters. These are small molecules synthesized in nerve cells that are packaged in membrane vesicles called synaptic vesicles in the nerve cell. Neurotransmitters come in all shapes and chemical forms, from small chemicals like acetylcholine to peptides like neuropeptide Y. The most abundant neurotransmitter is glutamate, which acts at over 90% of the synapses in the human brain.
Movie 3.2 - Movement of an action potential down a nerve cell - Wikipedia
Into the cleft
As the action potential in the presynaptic neuron approaches the axon terminus, synaptic vesicles begin to fuse with the membrane and their neurotransmitter contents spill into the synaptic cleft. Once in the cleft, the neurotransmitters diffuse, some of them reaching receptors on the postsynaptic cell. Binding of the neurotransmitter to the receptors on the membrane of the postsynaptic cell stimulates a response.
For motor neurons, the postsynaptic cell will be a muscle cell, and the response will be muscle contraction/relaxation. At this point, the originating nerve cell has done its job and communicated its information to its immediate target. If the postsynaptic cell is a nerve cell, the process repeats in that cell until it gets to its destination.
N
Distance Ed
To the tune of “Mister Ed”
Metabolic Melodies Website HERE
A course is a source,
of course, of course
Of all of the knowledge that we endorse
A major force for better/worse is the campus Distance Ed
It’s true to outsource a college course
There are a few standards to be enforced
The long and short’s we reinforce the campus Distance Ed
Bridge
A classroom class meets every week the same time every day
But Distance Ed is most unique - its flexible schedule’s okay
E-course is a source, of course, of course
Of online assistance for lab reports
You’re not enrolled in an online course?
Then sign up for this!
“You’ll love Distance Ed”
Recording by David Simmons
Lyrics by Kevin Ahern
Recording by David Simmons Lyrics by Kevin Ahern
313
It's one o'clock and
Ahern's talkin'
Henderson and
Hasselbalch and
pKa's and
Buffers I should know
This song's for BB three five oh
I hope that maybe
He'll think the way we
Wrote our answers
Wasn't crazy
I really need the
Partial credit - so
This song's for BB three five oh
It's really groovy
That it improves me
Watching lectures
In Quicktime movies
I really need to
Go and download those
Podcasts for BB three five oh
This Song's For BB 3-5-0
To the tune of "This Land is My Land"
Metabolic Melodies Website HERE
I'm feeling manic
I'm in a panic
I'd better study
My old organic
It has reactions
That I need to know
This song's for BB three five oh
I know he said it
That's why I dread it
'cause I skipped Friday's
Extra credit
'twil pro'bly haunt me
That lowly ze-ro
Grade in BB three five oh
It could be steric
Or esoteric
That carbons get so
Anomeric
I'm too hysteric
Better let it go
This song's for BB three five oh
Recording by Tim Karplus
Lyrics by Kevin Ahern
Recording by Tim Karplus Lyrics by Kevin Ahern