
5.2: Structure and Function - Lipids and Their Transport
- Page ID
- 354298

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Source: BiochemFFA_2_7.pdf. The entire textbook is available for free from the authors at http://biochem.science.oregonstate.edu/content/biochemistry-free-and-easy
Lipids are a diverse group of molecules that all share the characteristic that at least a portion of them is hydrophobic. Lipids play many roles in cells, including serving as energy storage (fats/oils), constituents of membranes (glycerophospholipids, sphingolipids, cholesterol), hormones (steroids), vitamins (fat soluble), oxygen/ electron carriers (heme), among others. For lipids that are very hydrophobic, such as fats/ oils, movement and storage in the aqueous environment of the body requires special structures. Other, amphipathic lipids, such as glycerophospholipids and sphingolipids spontaneously organize themselves into lipid bilayers when placed in water. Interestingly, major parts of many lipids can be derived from acetyl-CoA.
Fatty acids
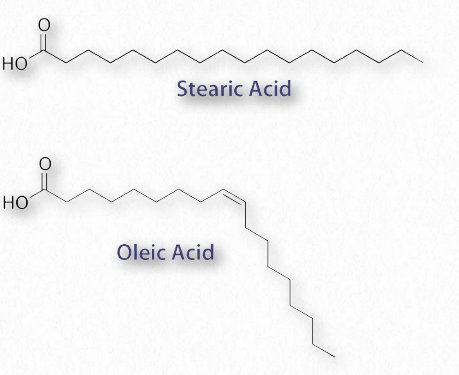
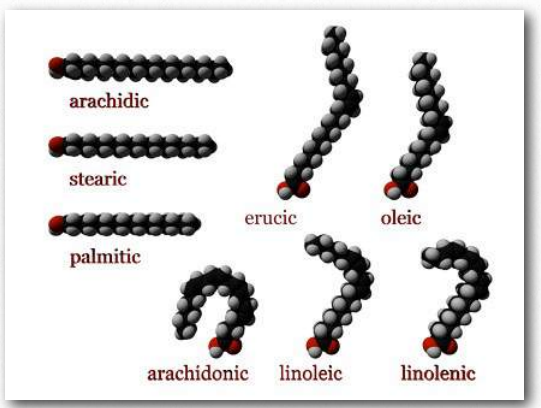
The most ubiquitous lipids in cells are the fatty acids. Found in fats, glycerophospholipids, sphingolipids and serving as as membrane anchors for proteins and other biomolecules, fatty acids are important for energy storage, membrane structure, and as precursors of most classes of lipids. Fatty acids, as can be seen from Figure 2.190 are characterized by a polar head group and a long hydrocarbon tail. Fatty acids with hydrocarbon tails that lack any double bonds are described as saturated, while those with one or more double bonds in their tails are known as unsaturated fatty acids. The effect of double bonds on the fatty acid tail is to introduce a kink, or bend, in the tail, as shown for oleic acid.
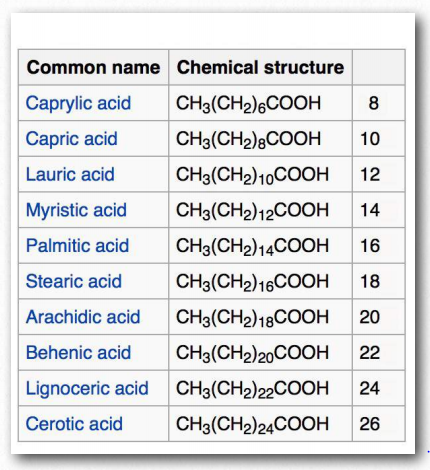
Stearic acid, a saturated fatty acid, by contrast has a straight hydrocarbon tail. Figures 2.190-2.194 show the most common saturated and unsaturated fatty acids. Fatty acids with unsaturated tails have a lower melting temperature than those with saturated tails of the same length. Shorter tails also decrease melting temperature. These properties carry over to the fats/oils containing them.
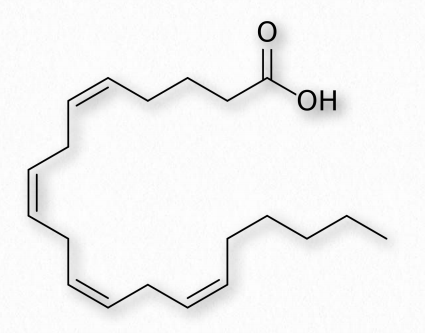
Fatty acids with more than one double bond are called polyunsaturated. Plants are excellent sources of unsaturated and polyunsaturated fatty acids. The position of the double bond(s) in fatty acids has important considerations both for their synthesis and for their actions in the body. Biochemically, the double bonds found in fatty acids are predominantly in the cis configuration. So-called trans fats arise as a chemical by-product of partial hydrogenation of vegetable oil.
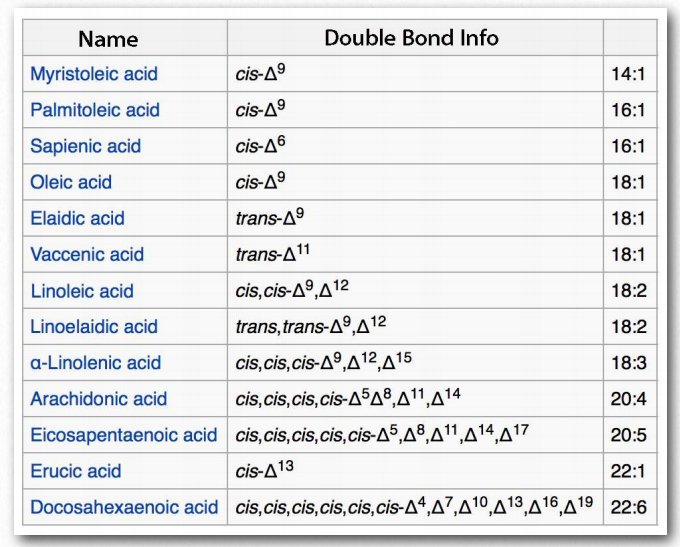
In humans, consumption of trans fat raises low density lipoprotein (LDL) levels and lowers high density lipoprotein (HDL) levels. Each is thought to contribute to the risk of developing coronary artery disease. The most Figure 2.194 - Fatty acid models. Carboxyl end labeled in red Wikipedia common fatty acids in our body include palmitate, stearate, oleate, linolenate, linoleate, and arachidonate. Two notable shorter fatty acids are nonanoic (9 carbons) and decanoic acid (10 carbons), both of which appear to have anti-seizure effects. Decanoic acid directly inhibits excitatory neurotransmission in the brain and may contribute to the anticonvulsant effect of the ketogenic diet.
Numbering
Figure 2.195 shows two different systems for locating double bonds in a fatty acid. The ω system counts carbons starting with the methyl end (shown in red) while the Δ system counts from the carboxyl end (shown in blue). For example, an ω-3 (omega 3) fatty acid would have a double bond at the third carbon from the methyl end. In the Δ system, a fatty acid that has a cis double bond at carbon 6, counting from the carboxyl end, would be written as cis-Δ6.
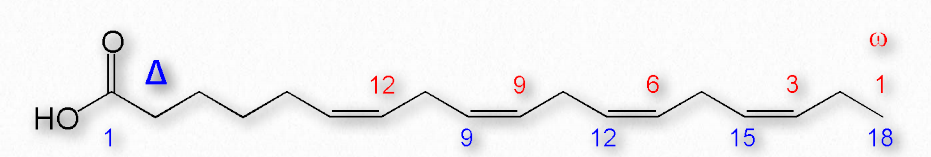
Fatty acids are described as essential fatty acids if they must be in the diet (can’t be synthesized by the organism) and nonessential fatty acids if the organism can synthesize them. Humans and other animals lack the desaturase enzymes necessary to make double bonds at positions greater than Δ-9, so fatty acids with double bonds beyond this position must be obtained in the diet. Linoleic acid and linolenic acid, both fall in this category. Related unsaturated fatty acids can be made from these fatty acids, so the presence of linoleic and linolenic acids in the diet eliminates the need to have all unsaturated fatty acids in the diet. Both linoleic and linolenic acid contain 18 carbons, but linoleic acid is an ω-6 fatty acid, whereas linolenic acid is an ω-3 fatty acid. Notably, ω-6 fatty acids tend to be proinflammatory, whereas ω-3 fatty acids are lesser so.

Fats/oils
Fats and oils are the primary energy storage forms of animals and are also known as triacylglycerols and triglycerides, since they consist of a glycerol molecule linked via ester bonds to three fatty acids (Figure 2.196). Fats and oils have the same basic structure. We give the name fat to those compounds that are solid at room temperature and the name oil to those that are liquid at room temperature. Note that biological oils are not the same as petroleum oils.
Increasing the number of unsaturated fatty acids (and the amount of unsaturation in a given fatty acid) in a fat decreases the melting temperature of it. Organisms like fish, which live in cool environments, have fats with more unsaturation and this is why fish oil contains polyunsaturated fatty acids.
Adipocytes
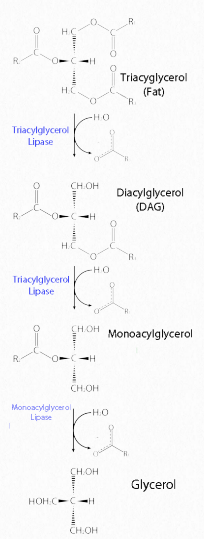
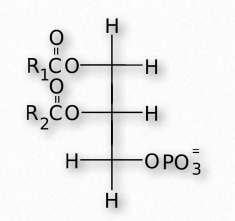
Fats are stored in the body in specialized cells known as adipocytes. Enzymes known as lipases release fatty acids from fats by hydrolysis reactions (Figure 2.197). Triacylglycercol lipase (pancreatic - Figure 2.198) is able to cleave the first two fatty acids from the fat. A second enzyme, monoacylglycerol lipase, cleaves the last fatty acid. Fats can be synthesized by replacing the phosphate on phosphatidic acid with a fatty acid.
Glycerophospholipids
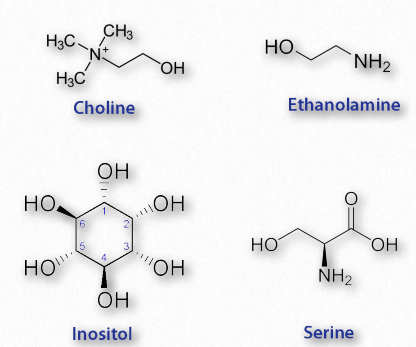
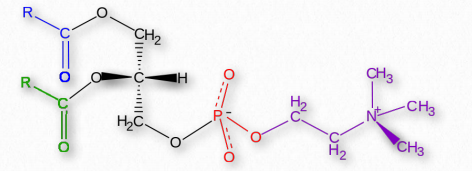
Glycerophospholipids (phosphoglycerides) are important components of the lipid bilayer of cellular membranes. Phosphoglycerides are structurally related to fats, as both are derived from phosphatidic acid (Figure 2.199). Phosphatidic acid is a simple glycerophospholipid that is usually converted into phosphatidyl compounds. These are made by esterifying various groups, such as ethanolamine, serine, choline, inositol, and others (Figure 2.200) to the phosphate of phosphatidic acid. All of these compounds form lipid bilayers in aqueous solution , due to their amphiphilic nature.
Phosphatidylethanolamines
Since all glycerolipids can have a variety of fatty acids at positions 1 and 2 on the glycerol, they all are families of compounds. The phosphatidylethanolamines are found in all living cells and are one of the most common phosphatides, making up about 25% of them. They are common constituents of brain tissue and in the spinal cord, making up as much as 45% of the total phospholipids. Phosphatidylethanolamines are asymmetrically distributed across membranes, being preferentially located on the inner leaflet (closest to the cytoplasm) of the plasma membrane. Metabolically, phosphatidylethanloamines are precursors of phosphatidylcholines. Phosphatidylserines Phosphatidylserines are another group of phosphatidyl compounds that are preferentially distributed across the lipid bilayer of the plasma membrane. Like the phosphatidylethanolamines, phosphatidylserines are preferentially located on the inner leaflet of the plasma membrane. When apoptosis (cell suicide) occurs, the preferential distribution is lost and the phosphatidylserines appear on the outer leaflet where they serve as a signal to macrophages to bind and destroy the cell.
Phosphatidylcholines
Phosphatidylcholines (Figure 2.201) are another group of important membrane components. They tend to be found more commonly on the outer leaflet of the plasma membrane. Nutritionally, the compounds are readily obtained from eggs and soybeans. Phosphatidylcholines are moved across membranes by Phosphatidylcholine transfer protein (PCTP). This protein, which is sensitive to the levels of phosphatidylcholines, acts to stimulate the activity of a thioesterase (breaks thioester bonds, such as acyl-CoAs) and activates PAX3 transcription factors.
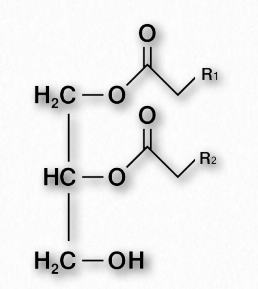
Diacylglycerol
Diacylglycerol (also called diglyceride and DAG - Figure 2.204) is an important intermediate in metabolic pathways. It is produced, for example, in the first step of the hydrolysis of fat and is also produced when membrane lipids, such as PIP2 (phosphatidylinositol-4,5-bisphosphate) are hydrolyzed by phospholipase C in a signaling cascade.
DAG is itself a signaling compound, binding to protein kinase C to activate it to phosphorylate substrates. Synthesis of DAG begins with glycerol-3-phosphate, which gains two fatty acids from two acyl-CoAs to form phosphatidic acid. Dephosphorylation of phosphatidic acid produces DAG. DAG can also be rephosphorylated by DAG kinase to re-make phosphatidic acid or another fatty acid can be added to make fat.
Signaling lipids
Inositol
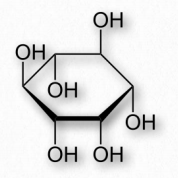
Though technically not a lipid itself, inositol is found in many lipids. Inositol is a derivative of cyclohexane containing six hydroxyl groups - one on each carbon (Figure 2.205. It has nine different stereoisomers of which one, cis-1,2,3,5-trans-4,6- cyclohexanehexol (called myo-inositol) is the most common. It has a sweet taste (half that of sucrose).
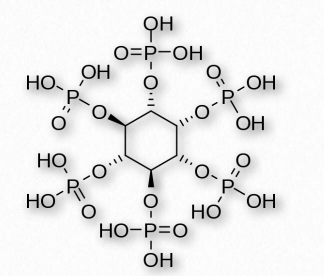
Numerous phosphorylated forms of the compound exist, from a single phosphate to six (one on each carbon). Phytic acid, for example, in plants, has six phosphates (Figure 2.206) that it uses to store phosphate. Inositol is produced from glucose and was once considered vitamin B8, but is made by the body in adequate amounts, so it is not now considered a vitamin. Phosphorylated forms of inositol are found in phosphoinositides, such as PIP2 and PIP3, both of which are important in signaling processes. Some of these include insulin signaling, fat catabolism, calcium regulation, and assembly of the cytoskeleton.
Phosphoinositides
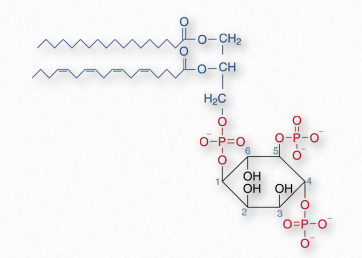
Compounds based on phosphatidylinositol (PI) are often called phosphoinositides. These compounds have important roles in signaling and membrane trafficking. Hydroxyls on carbons 3,4, and 5 of the inositol ring are targets for phosphorylation by a variety of kinases. Seven different combinations are used. Steric hindrance inhibits phosphorylation of carbons 2 or 6. Naming of these phosphorylated compounds follows generally as PI(#P)P, PI(#P, #P)P, or PI(#P, #P, #P)P where #P refers to the number of the carbon where a phosphate is located. For example, PI(3)P refers to a phosphatidyl compound with a phosphate added to carbons 3 of the inositol ring, whereas PI(3,4,5)P is a phosphatidyl compound with a phosphate added to carbons 3,4,and 5.
Phosphatidylinositol-4,5- bisphosphate
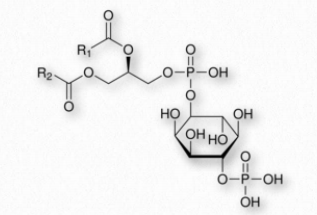
Phosphatidylinositol-4,5-bisphosphate (PIP2 - Figure 2.207) is a phospholipid of plasma membranes that functions in the phospholipase C signaling cascade. In this signaling pathway, hydrolysis catalyzed by phospholipase C releases inositol-1,4,5- trisphosphate (IP3) and diacylglycerol. Synthesis of PIP2 begins with phosphatidylinositol, which is phosphorylated at position 4 followed by phosphorylation at position 5 by specific kinases.
PIP2 can be phosphorylated to form the signaling molecule known as phosphatidylinositol (3,4,5)-trisphosphate (PIP3). Along with PIP3, PIP2 serves as a docking phospholipid for the recruitment of proteins that play roles in signaling cascades. Binding of PIP2 is also required by inwardly directed potassium channels.
Phosphatidylinositol (3,4,5)- trisphosphate
Phosphatidylinositol (3,4,5)-trisphosphate (PIP3) is an important molecule for the activation of signaling proteins, such as AKT, which activates anabolic signaling pathways related to growth and survival. PIP3 can be dephosphorylated by phosphatase PTEN to yield PIP2 and can be synthesized from PIP2 by kinase action of Class I PI 3- kinases. Kinase activity to synthesize PIP3 results in movement of PIP3-binding proteins to the plasma membrane. They include Akt/ PKB, PDK1, Btk1, and ARNO and each is activated by binding to PIP3.
Eicosanoids
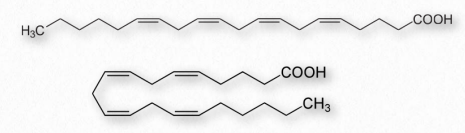
Fatty acids made from omega-6 and omega-3 fatty acids include three important fatty acids containing 20 carbons. They include arachidonic acid (an ω-6 fatty acid with four double bonds (Δ-5,8,11,14) - Figure 2.213), eicosapentaenoic acid (an ω-3 fatty acid with five double bonds, and dihomo-γ-linolenic acid (an ω-6 fatty acid with three double bonds). The class of compounds known as eicosanoids is made by oxidation of these compounds. Subclasses include include prostaglandins, prostacyclins, thromboxanes, lipoxins, leukotrienes, and endocannabinoids (Figures 2.214-2.219). Eicosanoids play important roles affecting inflammation, immunity, mood, and behavior.
Prostaglandins
A collection of molecules acting like hormones, prostaglandins are derived from arachidonic acid and have many differing (even conflicting) physiological effects. These include constriction or dilation of vascular smooth muscle cells, induction of labor, regulation of inflammation, and action on the thermoregulatory center of the hypothalamus to induce fever, among others.
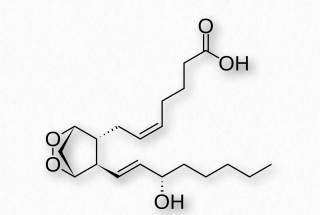
Prostaglandins are grouped with the thromboxanes (below) and prostacyclins (below), as prostanoids. The prostanoids, which all contain 20 carbons are a subclass of the eicosanoids. Prostaglandins are found in most tissues of higher organisms. They are autocrine or paracrine compounds produced from essential fatty acids. The primary precursor of the prostaglandins is the fatty acid known as arachidonic acid and the prostaglandin made from it is known as PGH2 (Figure 2.214), which, in turn is a precursor of other prostaglandins, as well as the prostacyclins and thromboxanes.
Interesting prostaglandins
PGD2 - inhibits hair follicle growth, vasodilator, causes bronchial constriction, higher in lungs of asthmatics than others.
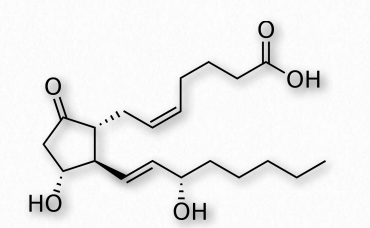
PGE2 (Figure 2.215) - exerts effects in labor (soften cervix, uterine contraction), stimulates bone resorption by osteoclasts, induces fever, suppresses T-cell receptor signaling, vasodilator, inhibits release of noradrenalin from sympathetic nerve terminals. It is a potent activator of the Wnt signaling pathway.
A prostaglandin can have opposite effects, depending on which receptor it binds to. Binding of PGE2 to the EP1 receptor causes bronchoconstriction and smooth muscle contraction, whereas binding of the same molecule to the EP2 receptor causes bronchodilation and smooth muscle relaxation.
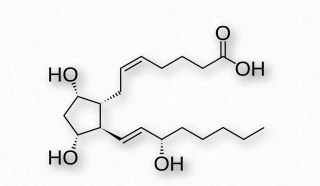
PGF2α (Figure 2.216)- uterine contractions, induces labor, bronchoconstriction.
PGI2 - vasodilation, bronchodilation, inhibition of platelet aggregation.
Thromboxanes
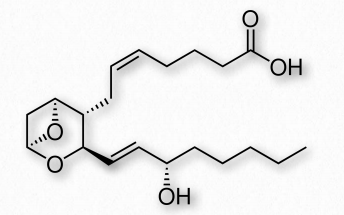
Thromboxanes play roles in clot formation and named for their role in thrombosis. They are potent vasoconstrictors and facilitate platelet aggregation. They are synthesized in platelets, as well. The anti-clotting effects of aspirin have their roots in the inhibition of synthesis of PGH2, which is the precursor of the thromboxanes. The most common thromboxanes are A2 (Figure 2.217) and B2.
Leukotrienes
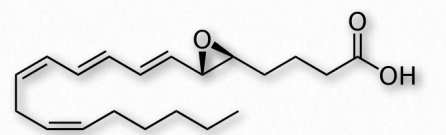
Another group of eicosanoid compounds are the leukotrienes (Figure 2.219). Like prostaglandins, leukotrienes are made from arachidonic acid. The enzyme catalyzing their formation is a dioxygenase known as arachidonate 5-lipoxygenase. Leukotrienes are involved in regulating immune responses. They are found in leukocytes and other immunocompetent cells, such as neutrophils, monocytes, mast cells, eosinophils, and basophils. Leukotrienes are associated with production of histamines and prostaglandins, which act as mediators of inflammation. Leukotrienes also trigger contractions in the smooth muscles of the bronchioles. When overproduced, they may pay a role in asthma and allergic reactions. Some treatments for asthma aim at inhibiting production or action of leukotrienes.
Steroids: membrane components and hormones
Steroids, such as cholesterol are found in membranes and act as signaling hormones in traveling through the body.
Cholesterol
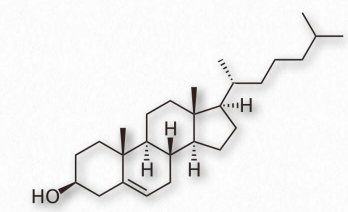
Arguably, no single biomolecule has generated as much discussion and interest as has cholesterol (Figure 2.220). Certainly, from the perspective of the Nobel Prize committee, no small molecule even comes close, with 13 people having been awarded prizes for work on it. Evidence for cholesterol’s importance comes from the study of brain tissue where it comprises 10-15% of the dry mass.
Membrane flexibility
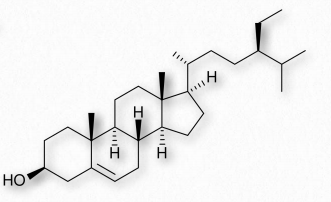
In animal cells, cholesterol provides for membrane flexibility that allows for cellular movement that is in contrast to plant and bacterial cells with fixed structures. Cholesterol is made in many cells of the body, with the liver making the greatest amount. The anabolic pathway leading to synthesis of cholesterol is known as the isoprenoid pathway and branches of it lead to other molecules including other fat-soluble vitamins.

Cholesterol is only rarely found in prokaryotes (Mycoplasma, which requires it for growth, is an exception) and is found in only trace amounts in plants. Instead, plants synthesize similar compounds called phytosterols (Figure 2.221). On average, the body of a 150 pound adult male makes about 1 gram of cholesterol per day, with a total content of about 35 grams.
Packaging
Cholesterol’s (and other lipids’) hydrophobicity requires special packaging into lipoprotein complexes (called chylomicrons, VLDLs, IDLs, LDLs, and HDLs) for movement in the lymph system and bloodstream. Though cholesterol can be made by cells, they also take it up from the blood supply by absorbing cholesterol-containing LDLs directly in a process called receptor-mediated endocytosis.
Oxidative damage to LDLs can lead to formation of atherosclerotic plaques and this is why cholesterol has gotten such a negative image in the public eye. The liver excretes cholesterol through the bile for elimination into the digestive system, but the compound is recycled there.
Reducing cholesterol levels
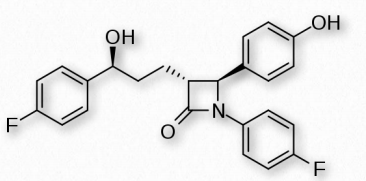
Strategies for reducing cholesterol in the body focus primarily on three areas - reducing consumption, reducing endogenous synthesis, and reducing the recycling. Dietary considerations, such as saturated fat versus unsaturated fat consumption are currently debated. Dietary trans fats, though, correlate with incidence of coronary heart disease. Consumption of vegetables may provide some assistance with reducing levels of cholesterol recycled in the digestive system, because plant phytosterols compete with cholesterol for reabsorption and when this happens, a greater percentage of cholesterol exits the body in the feces. Drugs related to penicillin are also used to inhibit cholesterol recycling. One of these is ezetimibe, shown in Figure 2.224.
Genetic defects in the cholesterol movement system are a cause of the rare disease known as familial hypercholesterolemia in which the blood of afflicted individuals contains dangerously high levels of LDLs. Left untreated, the disease is often fatal in the first 10-20 years of life. While LDLs have received (and deserve) a bad rap, another group of lipoprotein complexes known as the HDLs (high density lipoprotein complexes) are known as “good cholesterol” because their levels correlate with removal of debris (including cholesterol) from arteries and reduce inflammation.
Membrane function
In membranes, cholesterol is important as an insulator for the transmission of signals in nerve tissue and it helps to manage fluidity of membranes over a wide range of temperatures. Stacked in the lipid bilayer, cholesterol decreases a membrane’s fluidity and its permeability to neutral compounds, as well as protons and sodium ions. Cholesterol may play a role in signaling by helping with construction of lipid rafts within the cell membrane.
Steroid Hormones
Steroid hormones are all made from cholesterol and are grouped into five categories - mineralocorticoids (21 carbons), glucocorticoids (21 carbons), progestagens (21 carbons), androgens (19 carbons), and estrogens (18 carbons).
Mineralocorticoids
Mineralocorticoids are steroid hormones that influence water and electrolyte balances. Aldosterone (Figure 2.238) is the primary mineralocorticoid hormone, though other steroid hormones (including progesterone) have some functions like it. Aldosterone stimulates kidneys to reabsorb sodium, secrete potassium, and passively reabsorb water. These actions have the effect of increasing blood pressure and blood volume. Mineralocorticoids are produced by the zona glomerulosa of the cortex of the adrenal gland.
Glucocorticoids
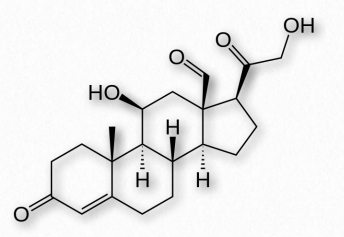
Glucocorticoids (GCs) bind to glucocorticoid receptors found in almost every vertebrate animal cell and act in a feedback mechanism in the immune system to reduce its activity. GCs are used to treat diseases associated with overactive immune systems. These include allergies, asthma, and autoimmune dis- Figure 2.237 - Steroid numbering scheme Image by Pehr Jacobson eases. Cortisol (Figure 2.239) is an important glucocorticoid with cardiovascular, metabolic, and immunologic functions. The synthetic glucocorticoid known as dexamethasone has medical applications for treating rheumatoid arthritis, bronchospasms (in asthma), and inflammation due to its increased potency (25-fold) compared to cortisol. Glucocorticoids are produced primarily in the zona fasciculata of the adrenal cortex.
Progestagens
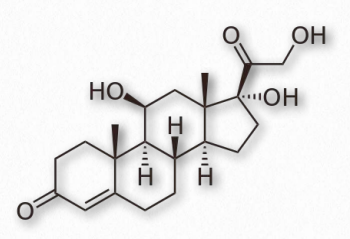
Progestagens (also called gestagens) are steroid hormones that work to activate the progesterone receptor upon binding to it. Synthetic progestagens are referred to as progestins. The most common progestagen is progesterone (also called P4 - Figure 2.240) and it has functions in maintaining pregnancy. Progesterone is produced primarily in the diestrus phase of the estrous cycle by the corpus luteum of mammalian ovaries. In pregnancy, the placenta takes over most progesterone production.
Androgens
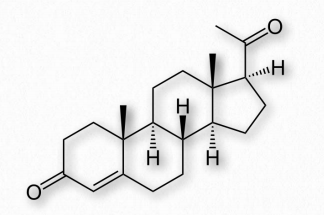
Androgens are steroid hormones that act by binding androgen receptors to stimulate development and maintenance of male characteristics in vertebrates. Androgens are precursors of estrogens (see below). The primary androgen is testosterone (Figure 2.241). Other important androgens include dihydrotestosterone (stimulates differentiation of penis, scrotum, and prostate in embryo) and androstenedione (common precursor of male and female hormones).
Estrogens
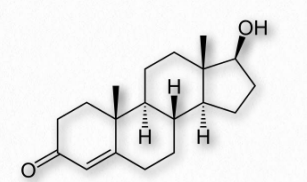
The estrogen steroid hormones are a class of compounds with important roles in menstrual and estrous cycles. They are the most important female sex hormones. Estrogens act by activating estrogen receptors inside of cells. These receptors, in turn, affect expression of many genes. The major estrogens in women include estrone (E1), estradiol (E2 - Figure 2.242), and estriol (E3). In the reproductive years, estradiol predominates. During pregnancy, estriol predominates and during menopause, estrone is the major estrogen.
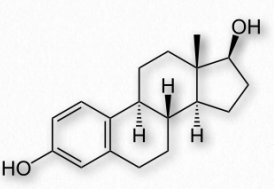
Estrogens are made from the androgen hormones testosterone and androstenedione in a reaction catalyzed by the enzyme known as aromatase. Inhibition of this enzyme with aromatase inhibitors, such as exemestane, is a strategy for stopping estrogen production. This may be part of a chemotherapeutic treatment when estrogenresponsive tumors are present.
Lipid Transport: lipoprotein complexes and lipid movement in the body
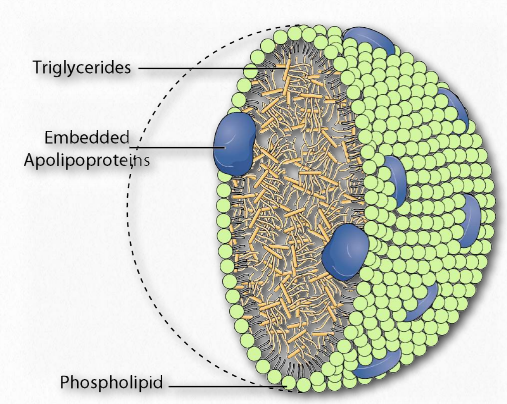
Lipoprotein complexes are combinations of apolipoproteins and lipids bound to them that solubilize fats and other non-polar molecules, such as cholesterol, so they can travel in the bloodstream between various tissues of the body. The apolipoproteins provide the emulsification necessary for this. Lipoprotein complexes are formed in tiny “balls” with the water soluble apolipoproteins on the outside and non-polar lipids, such as fats, cholesteryl esters, and fat soluble vitamins on the inside.
They are categorized by their densities. These include (from highest density to the lowest) high density lipoproteins (HDLs), Low Density Lipoproteins (LDLs), Intermediate Density Lipoproteins (IDLs), Very Low Density Lipoproteins (VLDLs) and the chylomicrons. These particles are synthesized in the liver and small intestines.
Apolipoproteins
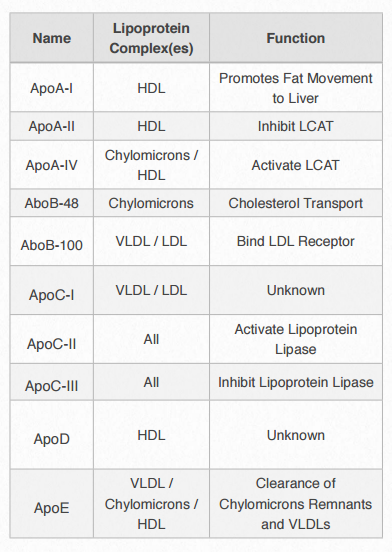
Each lipoprotein complex contain a characteristic set of apolipoproteins, as shown in Figure 2.256. ApoC-II and ApoC-III are notable for their presence in all the lipoprotein complexes and the roles they play in activating (ApoC-II) or inactivating (ApoC-III) lipoprotein lipase. Lipoprotein lipase is a cellular enzyme that catalyzes the breakdown of fat from the complexes. ApoE (see below) is useful for helping the predict the likelihood of the occurrence of Alzheimer's disease in a patient.
Deep dive: example of mRNA editing
ApoB-48 and ApoB-100 are interesting in being coded by the same gene, but a unique mRNA sequence editing event occurs that converts one into the other. ApoB-100 is made in the liver, but ApoB-48 is made in the small intestine. The small intestine contains an enzyme that deaminates the cytidine at nucleotide #2153 of the common mRNA. This changes it to a uridine and changes the codon it is in from CAA (codes for glutamine) to UAA (stop codon). The liver does not contain this enzyme and does not make the change in the mRNA. Consequently, a shorter protein is synthesized in the intestine (ApoB-48) than the one that is made in the liver (ApoB-100) using the same gene sequence in the DNA.
Movement
The movement of fats in the body is important because they are not stored in all cells. Only specialized cells called adipocytes store fat. There are three relevant pathways in the body for moving lipids. As described below, they are 1) the exogenous pathway (for fats that come in as food); 2) the endogenous pathway (for lipids that are synthesized in the body), and 3) the reverse transport pathway.
Exogenous pathway
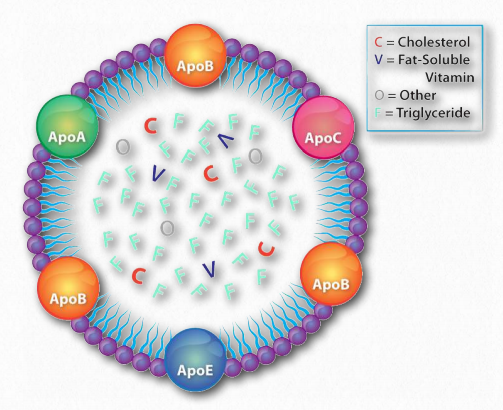
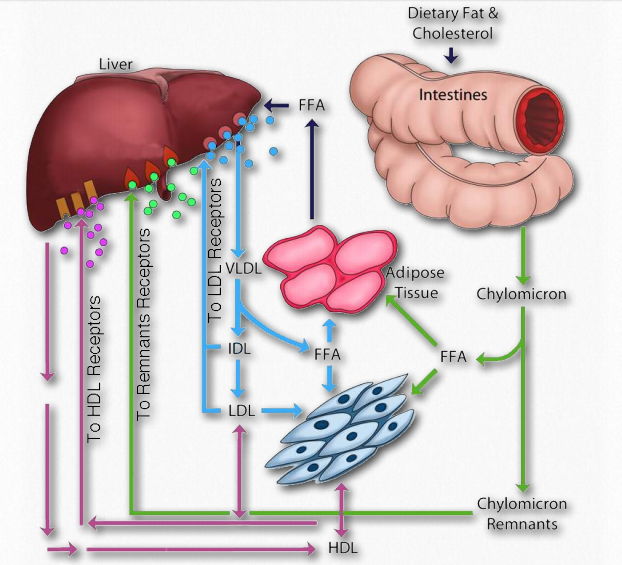
Dietary fat entering the body from the intestinal system must be transported, as appropriate, to places needing it or storing it. This is the function of the exogenous pathway of lipid movement in the body. All dietary lipids (fats, cholesterol, fat-soluble vitamins, and other lipids) are moved by it. In the case of dietary fat, it begins its journey after ingestion first by being solubilized by bile acids in the intestinal tract. After passing through the stomach, pancreatic lipases clip two fatty acids from the fat, leaving a monoacyl glycerol. The fatty acids and monoacyl glycerol are absorbed by intestinal cells (enterocytes) and reassembled back into a fat, and then this is mixed with phospholipids, cholesterol esters, and apolipoprotein B-48 and processed to form chylomicrons (Figures 2.258 & 2.259) in the Golgi apparatus and smooth endoplasmic reticulum.
Exocytosis
These are exocytosed from the cell into lymph capillaries called lacteals. The chylomicrons pass through the lacteals and enter the bloodstream via the left subclavian vein. Within the bloodstream, lipoprotein lipase breaks down the fats causing the chylomicron to shrink and become what is known as a chylomicron remnant. It retains its cholesterol and other lipid molecules.
The chylomicron remnants travel to the liver where they are absorbed (Figure 2.260). This is accomplished by receptors in the liver that recognize and bind to the ApoE of the chylomicrons. The bound complexes are then internalized by endocytosis, degraded in the lysosomes, and the cholesterol is disbursed in liver cells.
Endogenous pathway
The liver plays a central role in managing the body’s needs for lipids. When lipids are needed by the body or when the capacity of the liver to contain more lipids than is supplied by the diet, the liver packages up fats and cholesteryl esters into Very Low Density Lipoprotein (VLDL) complexes and exports them via the endogenous pathway. VLDL complexes contain ApoB-100, ApoC-I, ApoC-II, ApoC-III, and ApoE apolipoproteins. VLDLs enter the blood and travel to muscles and adipose tissue where lipoprotein lipase is activated by ApoC-II. In the muscle cells, the released fatty acids are taken up and oxidized. By contrast, in the adipoctyes, the fatty acids are taken up and reassembled back into triacylglycerides (fats) and stored in fat droplets. Removal of fat from the VLDLs causes them to shrink, first to Intermediate Density Lipoprotein (IDL) complexes (also called VLDL remnants) and then to Low Density Lipoprotein (LDL) complexes.
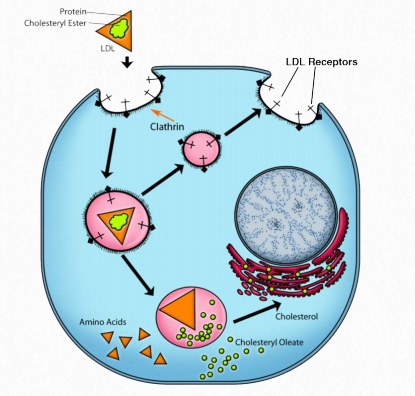
Shrinking of VLDLs is accompanied by loss of apolipoproteins so that LDLs are comprised primarily of ApoB-100. This lipoprotein complex is important because cells have receptors for it to bind and internalize it by receptor-mediated endocytosis (Figure 2.261). Up until this point, cholesterol and cholesteryl esters have traveled in chylomicrons, VLDLs, and IDLs as fat has been stripped stripped away. For cholesterol compounds to get into the cell from the lipoprotein complexes, they must be internalized by cells and that is the job of receptormediated endocytosis.
Reverse transport pathway
Another important consideration of the movement of lipids in the body is the reverse transport pathway (Figure 2.260). It is also called the reverse cholesterol transport pathway, since cholesterol is the primary molecule involved. This pathway involves the last class of lipoprotein complexes known as the High Density Lipoproteins (HDLs). In contrast to the LDLs, which are commonly referred to as “bad cholesterol” (see below also), the HDLs are known as “good cholesterol.”
HDLs are synthesized in the liver and small intestine. They contain little or no lipid when made (called depleted HDLs), but serve the role of “scavenger” for cholesterol in the blood and from remnants of other (damaged) lipoprotein complexes in the blood. To perform its task, HDLs carry the enzyme known as lecithincholesterol acyl transferase (LCAT), which they use to form cholesteryl esters using fatty acids from lecithin (phosphatidylcholine) and then they internalize them.
The cholesterol used for this purpose comes from the bloodstream, from macrophages, and from foam cells (macrophage-LDL complexes - Figure 2.262). Addition of cholesteryl esters causes the HDL to swell and Figure 2.261 - The process of receptor-mediated endocytosis Image by Aleia Kim when it is mature, it returns its load of cholesterol back to the liver or, alternatively, to LDL molecules for endocytosis. HDLs have the effect of lowering levels of cholesterol and it is for that reason they are described as “good cholesterol.”
Regulation of lipid transfer
It is important that cells get food when they need it so some control of the movement of nutrients is critical. The liver, which plays the central role in modulating blood glucose levels, is also important for performing the same role for lipids. It accomplishes this task the use of specialized LDL receptors on its surface. Liver LDL receptors bind LDLs that were not taken up by other cells in their path through the bloodstream. High levels of LDLs are a signal to the liver to reduce the creation of VLDLs for release.
People with the genetic disease known as familial hypercholesterolemia, which manifests with dangerously high levels of LDLs, lack properly functioning LDL receptors on their liver cells.Figure 2.263 - Progression of atherosclerosis Wikipedia Figure 2.263 - Progression of atherosclerosis Wikipedia Figure 2.263 - Progression of atherosclerosis Wikipedia
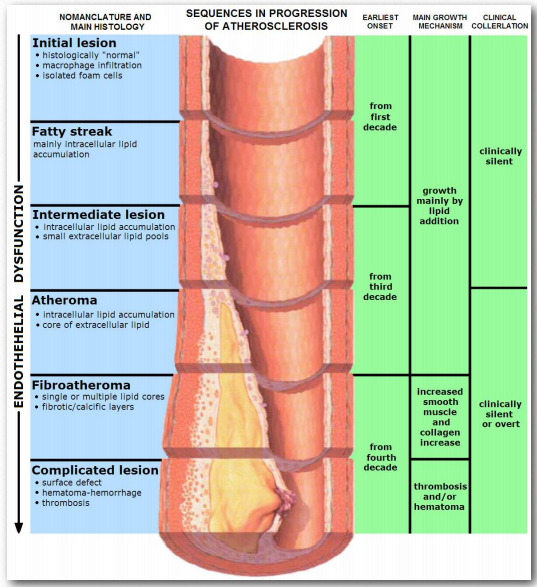
In sufferers of this disease, the liver never gets the signal that the LDL levels are high. In fact, to the liver, it appears that all VLDLs and LDLs are being taken up by peripheral tissues, so it creates more VLDLs to attempt to boost levels. Untreated, the disease used to be fatal early, but newer drugs like the statins have significantly increased life spans of patients. Cellular needs for the contents of LDLs are directly linked to the levels of synthesis of LDL receptors on their membranes. As cells are needing more cholesterol, their synthesis of components for receptors goes up and it decreases as need diminishes.
Good cholesterol / bad cholesterol
It is commonly accepted that “high cholesterol” levels are not healthy. This is due, at least indirectly, to the primary carriers of cholesterol, the LDLs. A primary function of the LDLs is to deliver cholesterol and other lipids directly into cells by receptor mediated endocytosis (Figure 2.237). High levels of LDLs, though, are correlated with formation of atherosclerotic plaques (Figure 2.263 & 2.264) and incidence of atherosclerosis, leading to the description of them as “bad cholesterol.” This is because when LDL levels are very high, plaque formation begins. It is thought that reactive oxygen species (higher in the blood of smokers) causes partial oxidation of fatty acid groups in the LDLs. When levels are high, they tend to accumulate in the extracellular matrix of the epithelial cells on the inside of the arteries. Macrophages of the immune system take up the damaged LDLs (including the cholesterol).
Since macrophages can’t control the amount of cholesterol they take up, cholesterol begins to accumulate in them and they take on appearance that leads to their being described as “foam cells.” With too much cholesterol, the foam cells, however, are doomed to die by the process of programmed cell death (apoptosis). Accumulation of these, along with scar tissue from inflammation result in formation of a plaque. Plaques can grow and block the flow of blood or pieces of them can break loose and plug smaller openings in the blood supply, ultimately leading to heart attack or stroke.
Good cholesterol
On the other hand, high levels of HDL are inversely correlated with atherosclerosis and arterial disease. Depleted HDLs are able to remove cholesterol from foam cells. This occurs as a result of contact between the ApoA-I protein of the HDL and a transport protein on the foam cell (ABC-G1). Another transport protein in the foam cell, ABCA-1 transports extra cholesterol from inside the cell to the plasma membrane where it is taken up into the HDL and returned to the liver or to LDLs by the reverse transport cholesterol pathway.
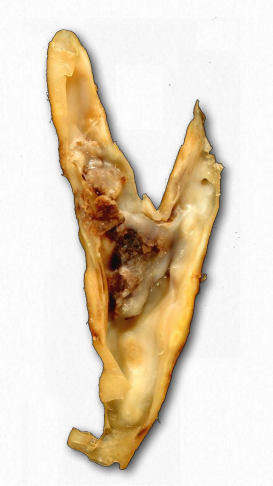
Deficiency of the ABCA-1 gene leads to Tangier disease. In this condition, HDLs are almost totally absent because they remain empty as a result of not being able to take up cholesterol from foam cells, so they are destroyed by the body.
ApoE and Alzheimer’s disease
ApoE is a component of the chylomicrons and is also found in brain, macrophages, kidneys, and the spleen. In humans, it is found in three different alleles, E2, E3, and E4. The E4 allele (present at about 14% of the population) is associated with increased likelihood of contracting Alzheimer's disease. People heterozygous for the allele are 3 times as likely to contract the disease and those homozygous for it are 15 times as likely to do so. It is not known why this gene or allele is linked to the disease. The three alleles differ only slightly in amino acid sequence, but the changes do cause notable structural differences. The E4 allele is associated with increased calcium ion levels and apoptosis after injury. Alzheimer’s disease is associated with accumulation of aggregates of the β- amyloid peptide. ApoE does enhance the proteolytic breakdown of it and the E4 isoform is not as efficient in these reactions as the other isoforms.