
Lipid Biosynthesis
- Page ID
- 5879

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)The complex organic compounds found in living organisms on this planet originate from photosynthesis, an endothermic reductive condensation of carbon dioxide requiring light energy and the pigment chlorophyll.
\[x CO_2 + x H_2O + \text{energy} \rightarrow C_xH_{2x}O_x + x O_2\]
The products of photosynthesis are a class of compounds called carbohydrates, the most common and important of which is glucose (C6H12O6). Subsequent reactions effect an oxidative cleavage of glucose to pyruvic acid (CH3COCO2H), and this in turn is transformed to the two-carbon building block, acetate. The multitude of lipid structures described here are constructed from acetate by enzymatic reactions that in many respects correspond to reactions used by chemists for laboratory syntheses of similar compounds. However, an important restriction is that the reagents and conditions must be compatible with the aqueous medium, neutral pH and moderate temperatures found in living cells. Consequently, the condensation, alkylation, oxidation and reduction reactions that accomplish the biosynthesis of lipids will not make use of the very strong bases, alkyl halides, chromate oxidants or metal hydride reducing agents that are employed in laboratory work.
Condensations
Claisen condensation of ethyl acetate (or other acetate esters) forms an acetoacetate ester, as illustrated by the top equation in the following diagram. Reduction, dehydration and further reduction of this product would yield an ester of butyric acid, the overall effect being the elongation of the acetate starting material by two carbons. In principle, repetition of this sequence would lead to longer chain acids, made up of an even number of carbon atoms. Since most of the common natural fatty acids have even numbers of carbon atoms, this is an attractive hypothesis for their biosynthesis.
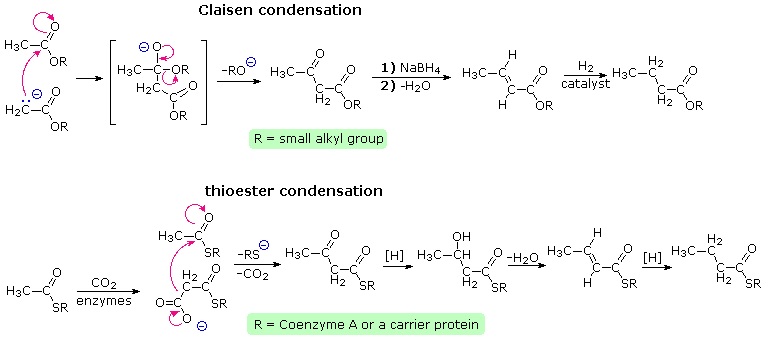
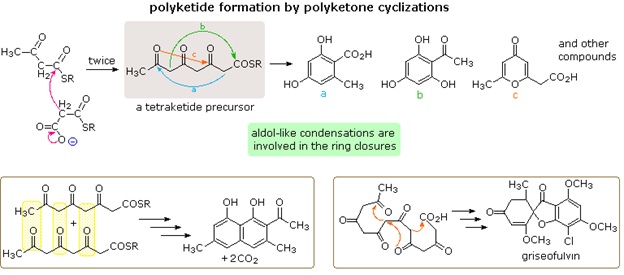
Nature's solution to carrying out a Claisen-like condensation in a living cell is shown in the bottom equation of the diagram. Thioesters are more reactive as acceptor reactants than are ordinary esters, and preliminary conversion of acetate to malonate increases the donor reactivity of this species. The thiol portion of the thioester is usually a protein of some kind, with efficient acetyl transport occurring by way of acetyl coenzyme A. Depending on the enzymes involved, the condensation product may be reduced and then further elongated so as to produce fatty acids (as shown), or elongated by further condensations to polyketone intermediates that are precursors to a variety of natural phenolic compounds.
The reduction steps (designated by [H] in the equations) and the intervening dehydrations needed for fatty acid synthesis require unique coenzymes and phosphorylating reagents. The pyridine ring of nicotinamide adenine dinucleotide (NAD) and its 2'-phosphate derivative (NADP) function as hydride acceptors, and the corresponding reduced species (NADH & NADPH) as a hydride donors. Partial structures for these important redox reagents are shown below.
As noted earlier, the hydroxyl group is a poor anionic leaving group (hydroxide anion is a strong base). Phosphorylation converts a hydroxyl group into a phosphate (PO4) or pyrophosphate (P2O7) ester, making it a much better leaving group (the pKas at pH near 7 are 7.2 and 6.6 respectively). The chief biological phosphorylation reagents are phosphate derivatives of adenosine (a ribose compound). The strongest of these is the triphosphate ATP, with the diphosphate and monophosphate being less powerful.
The overall process of fatty acid synthesis is summarized for palmitic acid, CH3(CH2)14CO2H, in the following equation:
\[8 CH_3CO-CoA + 14 NADPH + 14 H^+ + 7 ATP + H_2O \rightarrow CH_3(CH_2)_{14}CO_2H + 8 CoA + 14 NADP^+ + 7 ADP + 7 H_2PO_4^–\]
Alkylations
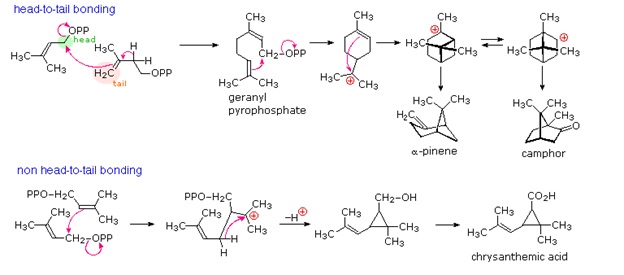
The branched chain and cyclic structures of the terpenes and steroids are constructed by sequential alkylation reactions of unsaturated isopentyl pyrophosphate units. As depicted in the following diagram, these 5-carbon reactants are made from three acetate units by way of an aldol-like addition of a malonate intermediate to acetoacetate. Selective hydrolysis and reduction gives a key intermediate called mevalonic acid. Phosphorylation and elimination of mevalonic acid then generate isopentenyl pyrophosphate, which is in equilibrium with its double bond isomer, dimethylallyl pyrophosphate. The allylic pyrophosphate group in the latter compound is reactive in enzymatically catalyzed alkylation reactions, such as the one drawn in the green box. This provides support for the empirical isoprene rule.
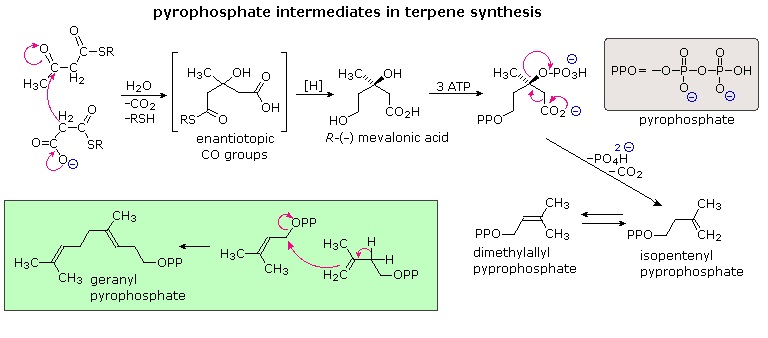
The simplest fashion in which isopentane units combine is termed "head-to-tail". This is the combination displayed in the green box, and these terms are further defined in the upper equation. Non head-to-tail coupling of isopentane units is also observed, as in the chrysanthemic acid construction shown in the second equation.
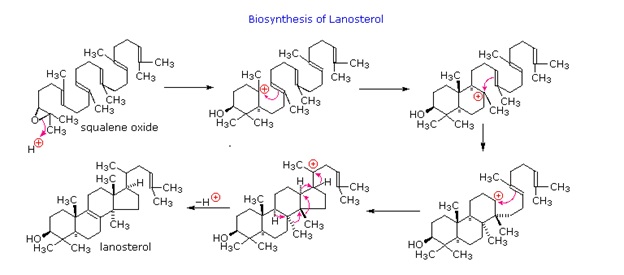
On the diagram below, the series of cation-like cyclizations and rearrangements, known as the Stork-Eschenmoser hypothesis, is shown, which were identified in the biosynthesis of the triterpene lanosterol. Lanosterol is a precursor in the biosynthesis of steroids. This takes place by metabolic removal of three methyl groups and degradation of the side chain.
Contributors
William Reusch, Professor Emeritus (Michigan State U.), Virtual Textbook of Organic Chemistry