
20.16: Higher-Order Structure
- Page ID
- 49626

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Several of the amino acid side chains are difficult to fit into either the α helix or β-sheet types of structure. An obvious example is proline, in which the R group is a ring and includes nitrogen bonded to the α carbon. When proline is involved in a peptide bond, all hydrogens are gone from the nitrogen, leaving no site for hydrogen bonding.
Another problem involves side chains having the same charge, such as those of lysine and arginine or glutamic acid and aspartic acid, which repel one another considerably. When they occur close together, these groups may also destabilize an α helix or β sheet structure. The presence of several groups of the type just mentioned allows a protein chain to bend sharply instead of laying flat or curling regularly into a spiral. Such sharp bends connect sections of α helix or β sheet structures in the globular proteins. They allow the polypeptide chain to curl back upon itself, folding the protein into a very compact, nearly spherical shape. The nature of this folding is referred to as tertiary structure, since it involves a third organizational level above the primary amino acid sequence and the secondary α helix or β sheet.
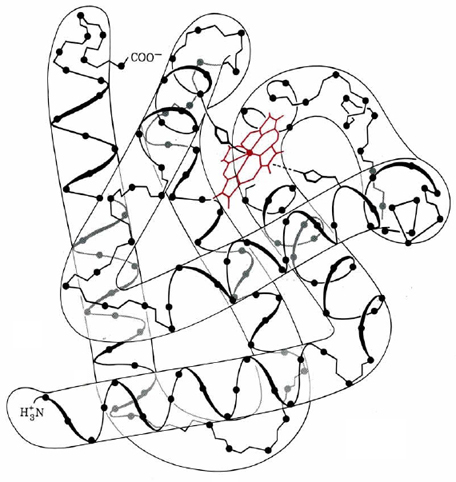
An excellent example of tertiary structure in a globular protein is provided by the three-dimensional view of sperm-whale myoglobin in Figure \(\PageIndex{1}\). The molecular backbone consists of eight relatively straight segments of a helix. The longest of these contain 23 amino acids, the shortest just 7. Fewer than 50 of myoglobin's 153 amino acids are found in the bends between the helixes, but all the proline residues fall into this category, as do many of the others which destabilize the α helix or β sheet. (Although only the α helix is prominent in the structure of myoglobin, other globular proteins are found to have regions consisting of β sheets.)
Several other important generalizations may be made about the structure of myoglobin. Even in regions where the chain twists into an α helix, nearly all the nonpolar R groups point toward the interior of the molecule. Here they crowd so closely together that only four water molecules can squeeze their way in. The outside surface of the protein, however, contains all the polar R groups. These interact strongly with the many water molecules which normally surround myoglobin in muscle tissue.
Although myoglobins isolated from a variety of mammals differ slightly in their primary structure, they all seem to have nearly the same overall molecular shape. Apparently some of the amino acids are much more important than others in determining the bend points and other crucial features of tertiary structure. Substitutions at less-important positions do not cause great variations in the ability of the protein to carry out its biological function.
Finally, Figure \(\PageIndex{1}\) shows clearly that myoglobin contains a prosthetic group. In this case it is the heme group, which contains an iron atom surrounded by four nitrogens in a flat ring structure known as a porphyrin. The porphyrin ring is not covalently bonded to the protein chain, but rather fits snugly into a “pocket” surrounded by several segments of helix. A nitrogen atom in a histidine side chain on one helix does form a coordinate covalent bond with iron in the heme group, but apart from this the prosthetic group is positioned solely by the way the protein chain is folded.
The iron in the heme group marks the active site at which the oxygen-storage function of myoglobin is accomplished. Iron, like other first-row transition-metal ions, ordinarily forms six bonds directed toward the corners of an octahedron. In myoglobin only five bonds to iron are found. The sixth position (opposite the histidine nitrogen) can be occupied by an oxygen molecule, providing a convenient storage site. If the concentration of oxygen near the protein falls, this binding is reversed, releasing oxygen to replenish the supply.
If an iron ion can bond to an oxygen molecule, you may wonder why the complicated porphyrin and protein structures are necessary. It is to prevent the oxidation of the iron in the heme group from the iron(II) to the iron(III) oxidation state. Because the oxygen molecule needs to gain two electrons to be oxidized while iron(II) can only supply one, no electron transfer occurs. If the iron(II) atom in heme were not surrounded by the protein chain, a water molecule would be able to take part as an intermediate in the electron-transfer mechanism and reduction of the oxygen could occur. As matters stand, though, the oxygen molecule cannot be reduced until it is released by the myoglobin. Thus the combined effects of the tertiary structure of the protein, the prosthetic group, and a specialized active site allow the myoglobin to fulfill its biological function of storing oxygen molecules until needed, without allowing them to be reduced during storage.
In some proteins there is yet a fourth level of organization, labeled quaternary structure. This may be illustrated by hemoglobin, whose function as an oxygen carrier in the bloodstream is well known. Hemoglobin consists essentially of four myoglobin molecules packed together in a single unit. Four separate polypeptide chains are each folded as in myoglobin and then nested together. The way these four subunits fit together constitutes the quaternary structure.
Like the other type of structures, quaternary structure contributes to the function of a protein. In the case of hemoglobin, an oxygen molecule attached to one subunit causes slight shifts in tertiary and quaternary structure which make it easier for other oxygen molecules to bond to the other subunits. Consequently hemoglobin in the lungs can be loaded with its full complement of four oxygen molecules rather easily, a factor which increases its efficiency in carrying oxygen to body tissues. The converse is also true—loss of one oxygen molecule causes slight structural rearrangements which allow the remaining three to depart more readily.