
1.4: Basic Metabolic Pathways
- Page ID
- 306241

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Basic Metabolism Overview
- Glycolysis (cytosol) and TCA cycle (mitochondria) convert glucose to high energy molecules: ________ and ___________ and __________.
This is just the beginning of energy production. NADH and FADH2 can be converted to more ATP. Oxidative phosphorylation is a metabolic pathway that transfers energy from NADH to the synthesis of ATP in the mitochondria.
- NADH oxidation occurs over many steps. Why don’t cells do this reaction directly? (Hint: This is a hydride reaction!)
Cellular Locations
Electrons stored in the form of the reduced coenzymes, NADH or FADH2, are passed through a chain of proteins and coenzymes to reduce O2 – the terminal electron acceptor – into H2O.
- NADH is formed at what point in metabolism: ___________.
- The TCA cycle occurs in __________.
- This electron transfer of oxidative phosphorylation occurs in ________________.
ATP production
The energy released by electrons flowing through this electron transport chain is used to transport protons to generate a pH gradient across the membrane.
- The phosphorylation of ADP to form ATP is [endothermic or exothermic].
- Protons to flow back across the membrane to restore equilibrium. This process is [diffusion or active transport ] and can drive a reaction.
Basic Metabolism: Glycolysis
Glucose is metabolized to produce energy (ATP) for the cell with the release of CO2 and H2O as byproducts. Glycolysis is a series of enzyme-catalyzed reactions that break glucose into 2 equivalents of pyruvate. This process (summarized below) is also called the Embden-Meyerhoff pathway.
- How many ATP are produced in this process? Keep in mind that everything is doubled after the 6 C glucose is cleaved into 2 3C units.
- How many ATP are consumed?
- Glycolysis results in the net formation of:
- ______ NADH
- ______ ATP
- ______ H2O
- Is glycolysis an uphill or downhill process? (i.e. exothermic or endothermic?)
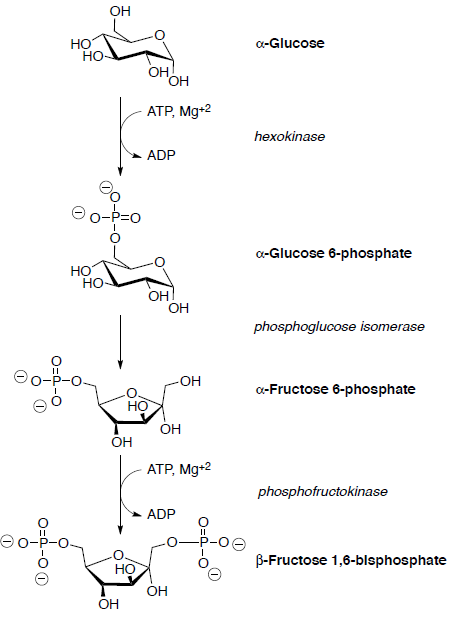
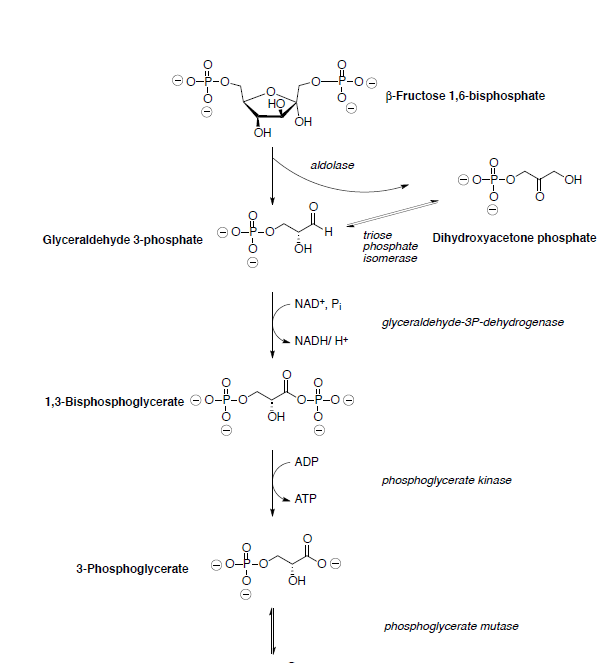
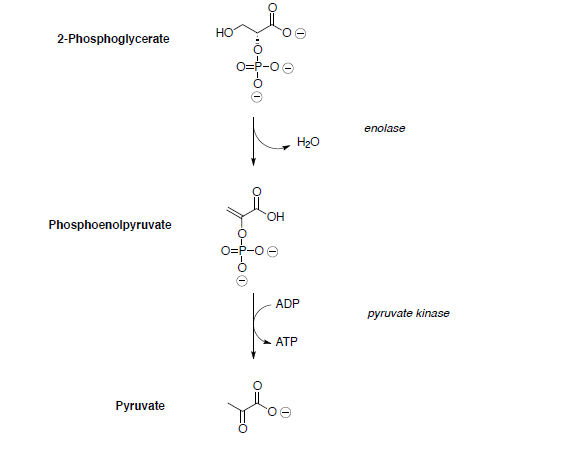
Assume all reactions take place within an enzyme.
Glucose is first phosphorylated at the hydroxyl group on C6 by reaction with ATP.
- Propose a mechanism for this reaction.
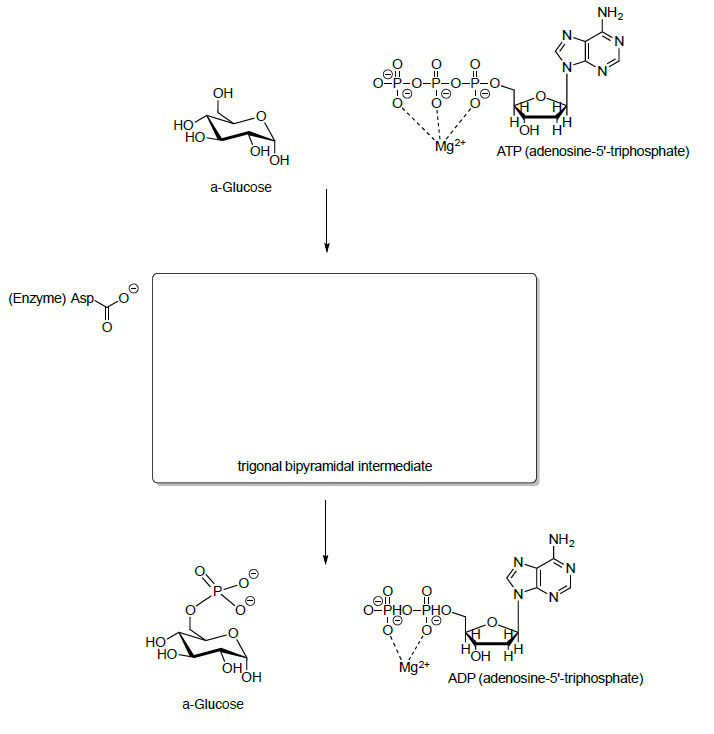
- ATP is not that reactive on its own. Why?
- Explain why a phosphate ester is a good electrophile when the Mg+2 is around.
Glucose-6-phosphate is isomerized to fructose-6-phosphate in the next step. The glucose-fructose interconversion is a multistep process whose details are not yet fully understood.
It begins with opening of the hemiacetal to an open-chain aldehyde.
- Propose a mechanism for this reaction.
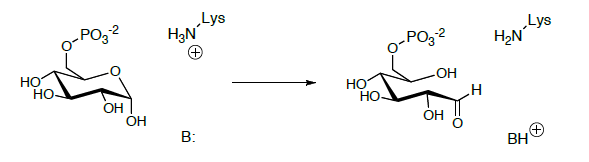
The open-chain aldehyde undergoes keto-enol tautomerization to the enediol which is further tautomerized to a different keto form.
- Looking at the structures of the sugars, propose a mechanism for this reaction.
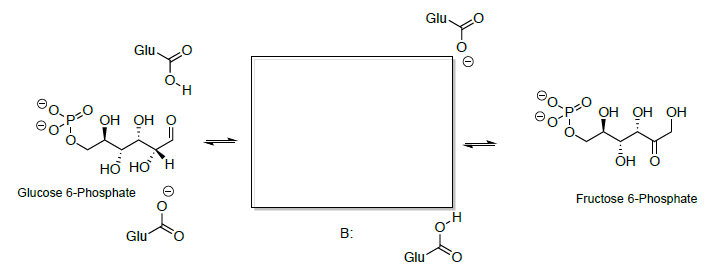
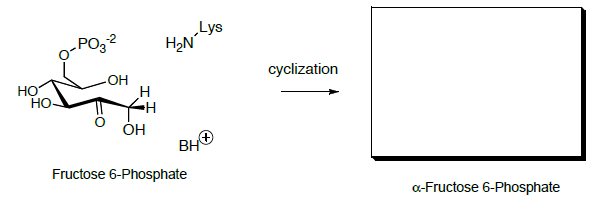
Cyclization of the open-chain hydroxy ketone gives fructose (hemiacetal).
- Show a mechanism.
- Predict the product.

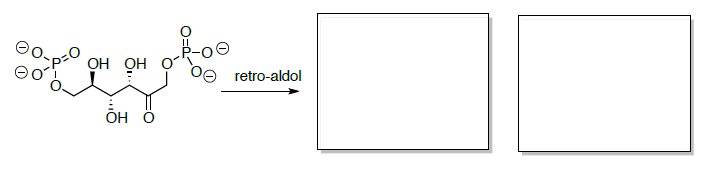
Fructose-6-phosphate is then converted to fructose 1,6-bisphosphate which is subsequently cleaved into two three-carbon compounds through a retro-aldol.
Review: aldol reaction
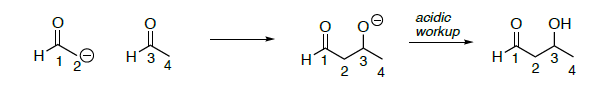
- On the aldol reaction above,
- Put a circle around the nucleophile
- Put a box around the electrophile in your starting materials
- Highlight the bond that is formed (broken in the retro reaction)
Retro-Aldol
If the reaction is driven to starting materials (retro-aldol), then the reaction will favor the starting materials.
- Draw the mechanism for the retro-aldol when starting with fructose 1,6-bisphosphate.
- Predict the two products of this retro aldol reaction.
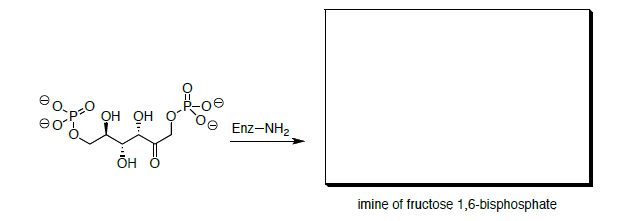
This mechanism is actually completed with an imine. Fructose 1,6-bisphosphate first reacts with the amino group of a lysine residue from an enzyme.
- Draw a mechanism for the formation of the imine.
The imine can then do a ‘retro-Stork enamine’ reaction (similar to the retro-aldol).
Review: Stork Enamine (an adol with the enamine replacing the enolate anion as the nucleophile).
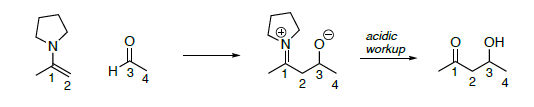
- On the enamine reaction above,
- Put a circle around the nucleophile
- Put a box around the electrophile in your starting materials
- Highlight the bond that is formed (broken in the retro reaction)
Retro-Stork enamine
If the reaction is driven to starting materials (retro-Stork enamine), then the reaction will favor the enamine and aldol starting materials.
- Predict the two products formed.
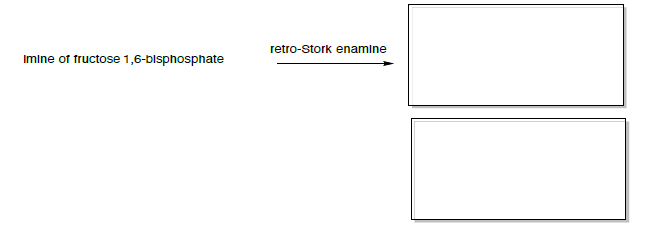
The products of the retro-Stork enamine are the enamine of dihydroxyacetone phosphate and glyceraldehyde 3-phosphate (shown below).
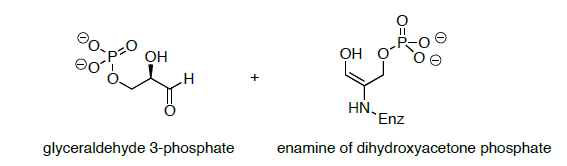
- Propose a mechanism for the conversion of the enamine of dihydroxyacetone phosphate is converted to a second molecule of glyceraldehyde 3-phosphate.

Glyceraldehyde 3-phosphate is oxidized and phosphorylated to 1,3-bisphosphoglycerate.
- Show the mechanisms for this transformation.
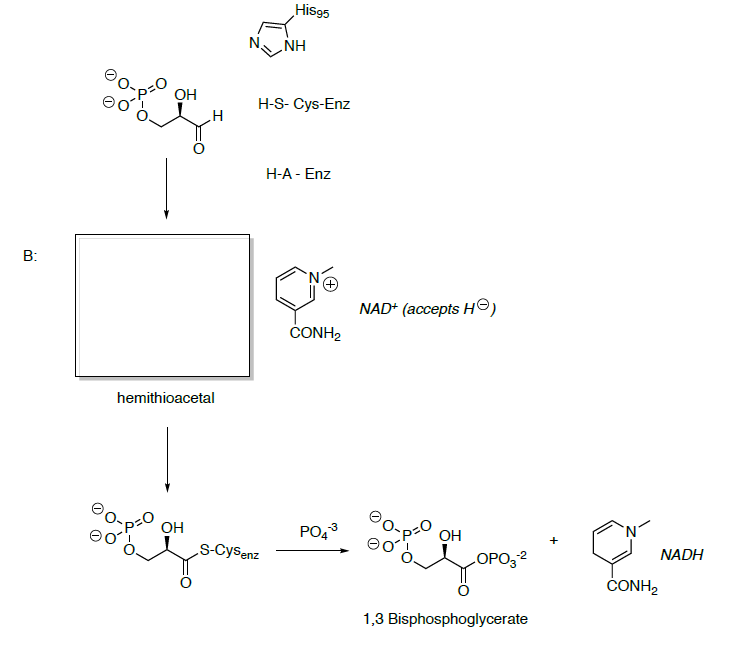
- What is the functional group formed in 1,3-bisphosphoglycerate?
- Predict the reactivity of this carbonyl.
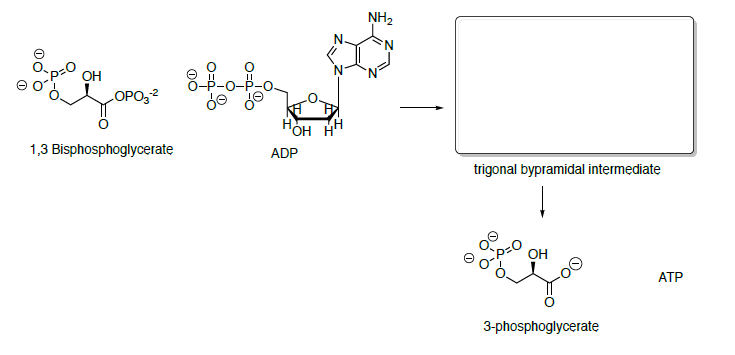
Phosphoglycerate kinase catalyzes the transfer of a phosphoryl group from 1,3-bisphosphoglycerate to ADP forming ATP and 3-phosphoglycerate.
- Propose a mechanism for this transformation.
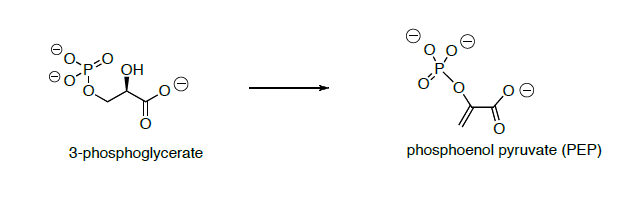
3-phosphoglycerate is converted to phosphoenol pyruvate (PEP) through dehydration and dephosphorylation.
In the last step of the metabolic breakdown of sugars (glycolysis), an enol phosphate is converted to pyruvic acid (shown below). The pyruvic acid is then converted to Acetyl Co A, which is the beginning of the TCA cycle.
- Draw a mechanism for the conversion of the enol phosphate to pyruvic acid.
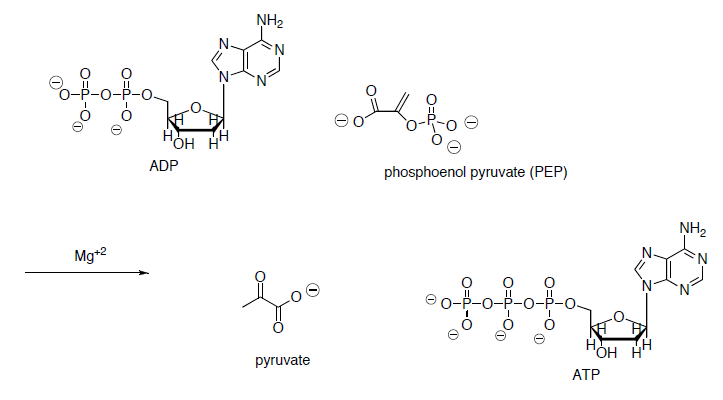
- What drives this reaction? (ie what factors make this reaction energetically favorable?)
Basic Metabolism: TCA Cycle
Hans Krebs and Fritz Lipmann shared the Nobel Prize for Physiology and Medicine in 1953 for their work on elucidating the Krebs cycle and coenzyme A. The Krebs Cycle [or tricarboxylic acid (TCA) or citric acid cycle] plays a central role in the metabolism of glucose to produce energy (ATP). The TCA cycle results ultimately in the oxidation of acetic acid to two molecules of carbon dioxide.
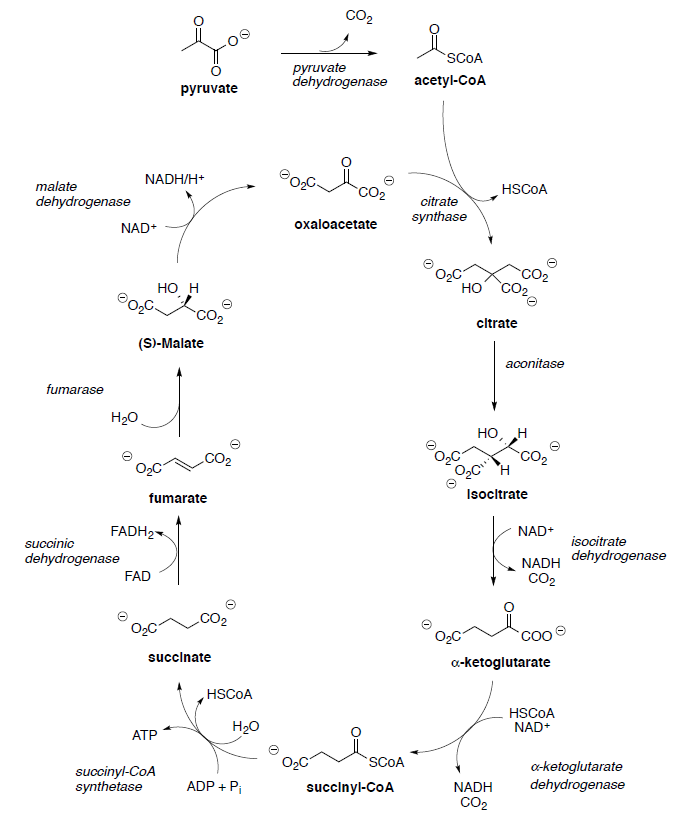
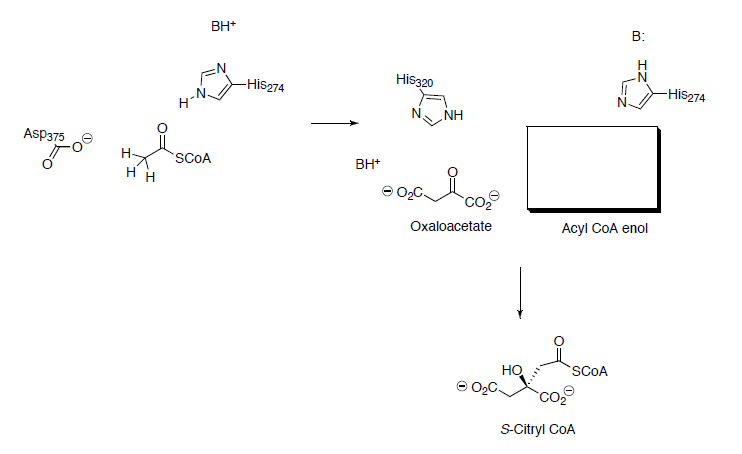
Pyruvate (end product of glycolysis) must be converted to acetyl CoA to enter the TCA cycle.
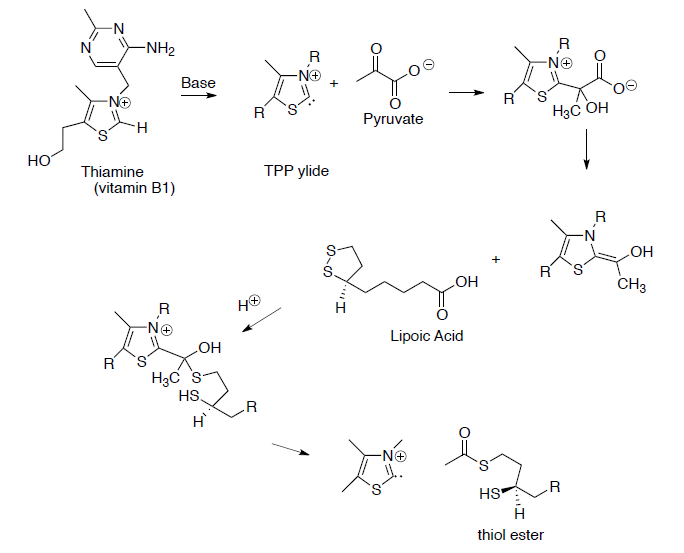
This process begins with the formation of a thiol ester from pyruvate.
- Draw reaction mechanisms for the steps shown below.
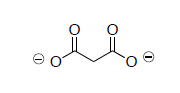
- In his experiments that led to the elucidation of the TCA cycle, Hans Krebs added malonate (shown below) to extracts of pigeon flight muscle. The malonate could not be used as a substrate to replace pyruvate in the pathway above. Why can’t malonate be used? (Think of the carbonyl hill).
At this point, co-enzyme A reacts with the thiol ester (formed in question on previous page) to form acetyl CoA (shown below). To help keep track of the sulfurs, one is in a box and one is in a circle.
- Draw the mechanism for this reaction.
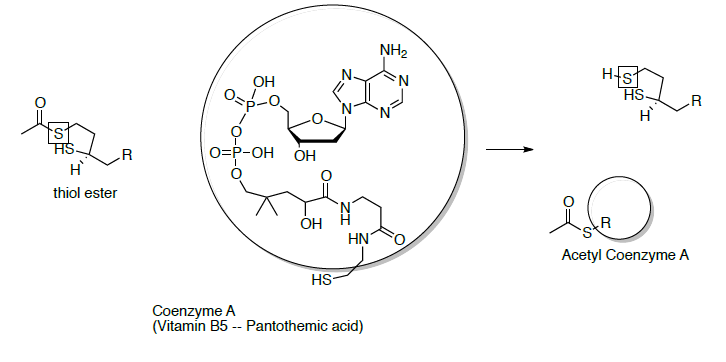
- The thiol ester formed in the last step of the reaction above is an ‘activated carbonyl’ (i.e. a better electrophile). Explain why the thiol ester is a better electrophile than the carboxylate anion.
- In an equivalent organic chemistry reaction, what would you use as an ‘activated carbonyl’?
In the next step Acetyl CoA reacts with oxaloacetate to form citryl CoA.
- Propose a mechanism for this reaction.
- In a similar reaction in organic chemistry, what would be the product for the reaction below? What type of reaction is this?
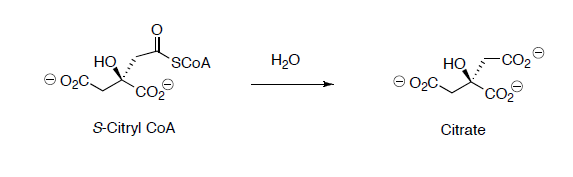
Citryl CoA is then hydrolyzed to citrate.
- Propose a reaction mechanism for this reaction.
Citrate is converted to isocitrate through two steps.
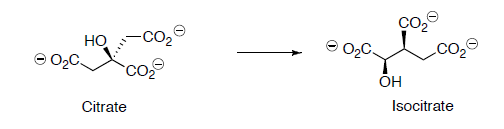
- Label all chiral centers with R or S.
- What changed in the conversion of citrate to isocitrate?
Isocitrate is oxidized to oxalosuccinate with NAD+.
- Draw the mechanism (and the other product) for this reaction.
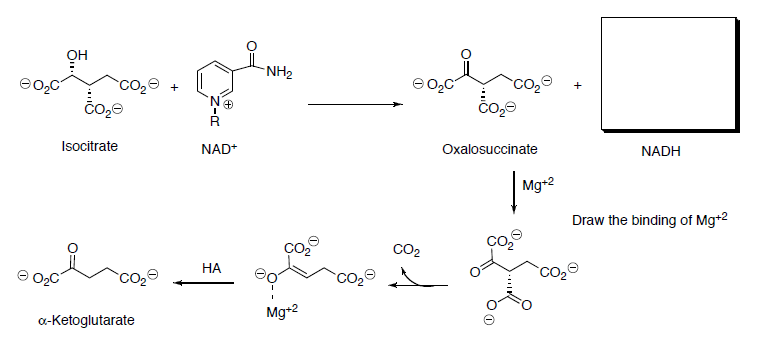
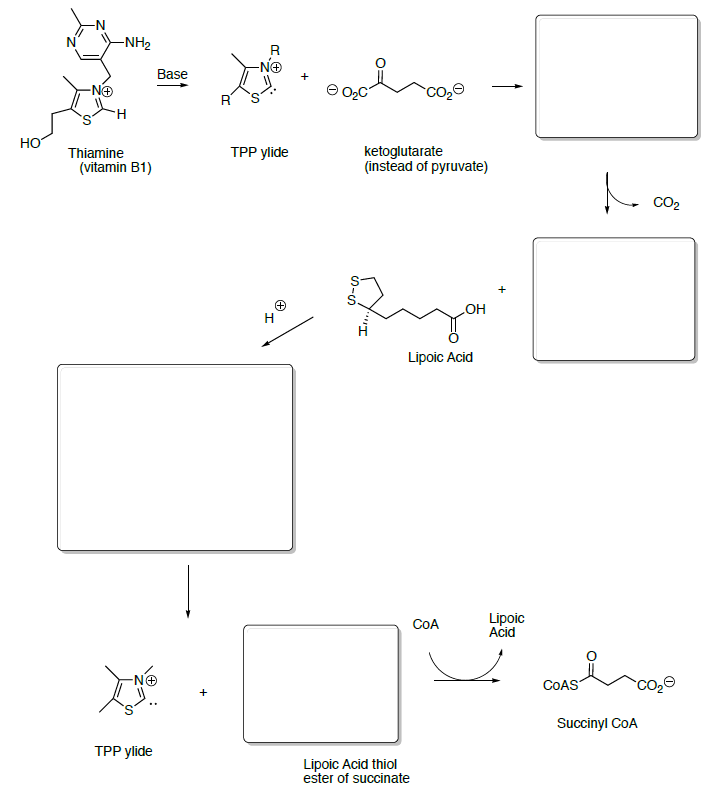
Ketoglutarate is transformed to succinyl CoA in a multistep process analogous to the transformation of pyruvate to acetyl CoA that we saw in the first step.
- Draw the transformation starting with the reaction with TPP ylide.
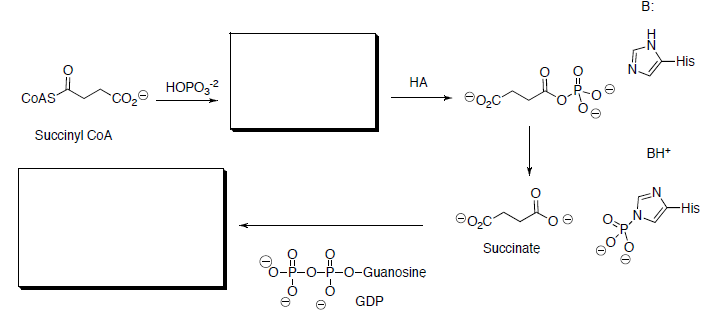
Succinyl CoA is hydrolyzed to succinate and is coupled with the phosphorylation of guanosine diphosphate (GDP) to give guanosine triphosphate (GTP).
- Draw the mechanism for this reaction.
Basic Metabolism: Oxidative Phosphorylation
Electron Transfer in Complex I
Complex I is located in the inner mitochondrial membrane in eukaryotes. The electrons from NADH (produced in the TCA cycle) begin to be shuttled through small steps to capture the energy.
This section will examine the mechanisms of electron transfer by the peripheral domain, proton transfer by the membrane domain and how their coupling can drive proton transport.
The net reaction of Complex I is the oxidation of NADH and the reduction of ubiquinone.
Net reaction:
\[\ce{NADH + H^+ + UQ \rightarrow NAD^+ + UQH2}\]
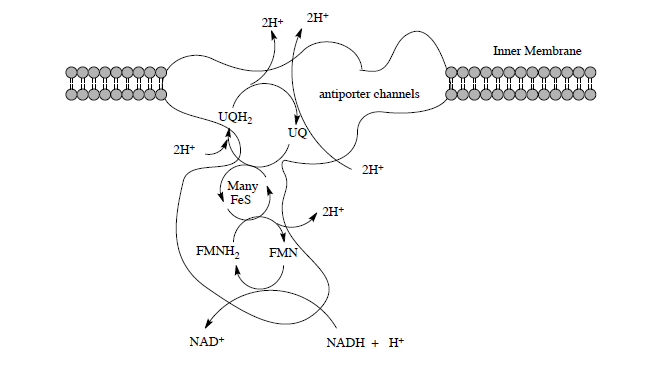
- How many protons are moved across the membrane for each cycle of Complex I?
- Is this active transport or passive diffusion?
- If this is active, what is fueling this transport?
- Is this with or against the concentration gradient? (i.e. antiporter or synporter?)
Complex II: Overview
Complex II (aka succinate dehydrogenase from the TCA cycle) oxidizes succinate (–O2CCH2CH2CO2–) to fumarate (trans-–O2CCH=CHCO2–).
Complex II also has a cascade of electron transfers. When succinate is converted to fumarate, the electrons are passed through a new cascade to eventually reduce UQ (just like Complex I!)
\[\ce{succinate \rightarrow fumarate + 2H+ + 2e-}\]
\[\ce{UQ + 2H+ + 2e- \rightarrow UQH2}\]
- Write the net reaction for the work of Complex II.
- The reaction catalyzed in Complex II has a very small ΔG°. Is it sufficient to power an antiporter channel?
Complex III: Overview
Complex III (sometimes called cytochrome bc1 complex) has two main substrates: cytochrome c and UQH2. The structure of this complex was determined by Johann Deisenhofer (Nobel Prize for a photosynthetic reaction center – we will see this soon).
This role of complex III is to transfer the electrons from UQH2 to cytochrome c.
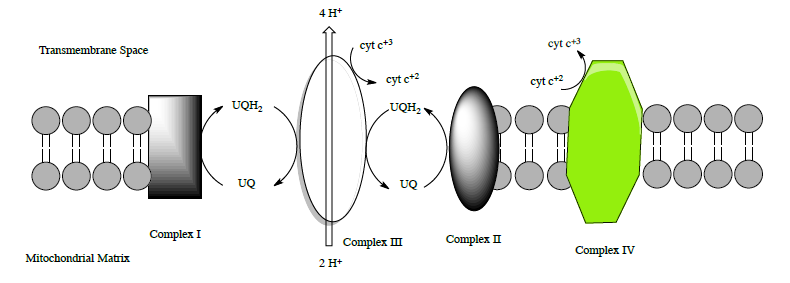
- Complete the equation for the redox reactions of complex III.
___ UQH2 + 1 UQ + 2 H+ + ___ cyt c+3 \(\ce{\rightarrow}\) ___ UQH2 + ___ UQ + 4 H+ + ___ cyt c+2
• There are two H+ coming from the mitochondrial matrix but _____ H+ are transported into the
inter-membrane space
Complex III to Complex IV: Cytochrome C as a mobile carrier
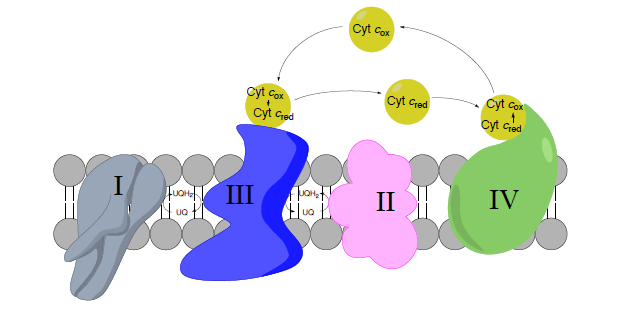
- Circle the mobile electron carriers in the picture above.
Complex IV Overview
Another complex whose goal is to move electrons and protons! This is the big step since it is the main site for dioxygen
utilization in all anaerobic organisms. The structure of complex IV is shown in the left figure and to the right in a diagram taken from the Kegg pathways (with permission).
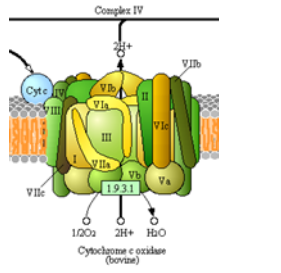
- Complete the net equation for the redox reactions of complex IV.
___ cyt c+2 + 1 O2 + 8 H+ \(\ce{\rightarrow}\) ___ H2O + 4 H+ + ___ cyt c+3
- How many protons are being “pumped” into the intermembrane space? _________
- How many electrons are needed to balance this equation? _________
- What are the initial and final “mobile” carriers of electrons?
Complex V: ATP Synthase
Neglecting Complex II, the overall reaction of the mitochondrial chain, per 2e– transferred, can be written as:
\[\ce{NADH + H+ + ½ O2 + 10 H+("in") \rightarrow NAD+ + H2O + 10 H+("out")} E° = +1.135V\]
Each two e– (from 1 NADH molecule) through the electron transport chain results in the net transfer of 10 protons across the membrane:
- Complex I: ________ H+
- Complex III: ________ H+
- Complex IV: ________ H+
Protons will diffuse from an area of high proton concentration to an area of lower proton concentration. Peter Mitchell received the Nobel Prize in 1978 for his proposal that an
electrochemical concentration gradient of protons across a membrane could be harnessed to make ATP. The proton gradient created by the electron transport chain provides enough energy to synthesize about 2.5 molecules of ATP through a process called chemiosmosis.
- This proton flow is driven by two forces (fill in the blanks):
- Diffusion force caused by a concentration gradient. All particles tend to move from __________ concentration to __________ concentration.
- Electrostatic force caused by an electrical potential gradient. An electrical gradient is a consequence of charge separation. Protons will be attracted to ___________.
ATP synthase is an important enzyme that utilizes the proton gradient drive the synthesis of (ATP).
Electric Potential Drives Motor
The rotor is not locked in a fixed position in the center of the bilayer and the rotor sites switch between the empty and the ion bound states. When driving ATP synthesis, an ion arrives from the periplasm and binds at an empty rotor site.
The positive stator charge (Arg227) plays a fundamental role in the function of the F0 motor.
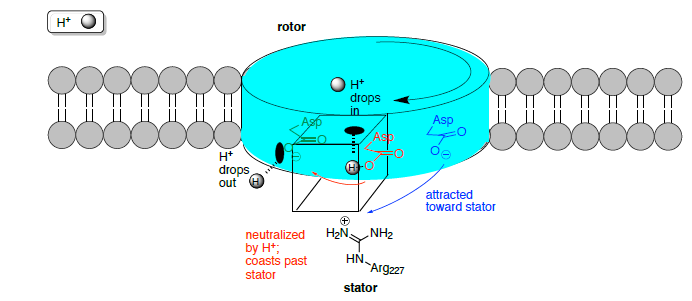
- What is the charge of the empty binding sites:
- when no ion is bound?
- when a Na+/H+ ion is binding?
- When an ion enters the rotor site from the stator channel, the net charge is reduced, thus [ increasing /decreasing ] the attraction to the stator. Now the rotor is able to move through the hydrophobic part of the stator, while the arginine attracts the next empty rotor site.
- The empty site (charge = ________) is electrostatically attracted by the stator (charge = ________) and guided into the next slot.
- The rotor site is occupied until it reaches the stator from the opposite side, where it encounters the positive stator charge, causing dissociation of the ion. Why? Consider diffusion gradients and charges.
Electrical Power Fuels Rotary ATP Synthase
- Fill in the blanks on the following summary of ATP Synthase:
During ATP synthesis, the ____________ gradient fuels the membrane-embedded F0 motor to rotate the central stalk. This rotation causes sequential binding changes at the peripheral F1 domain so that one catalytic site binds ________ and phosphate, the second makes tightly bound ATP, and the third step ____________.
In anaerobically growing bacteria, when the respiratory enzymes are not active, the F1 motor can hydrolyze ATP.
- Which direction will the pump turn in these conditions?
- What will happen to the F0 motor? And the H+ gradient?
Sources
Dimroth, Operation of the F0 motor of the ATP synthase, Biochimica et Biophysica Acta (BBA) -Bioenergetics, 2000, 1458, 374-386.