
10.8: Nucleophilic Carbonyl Addition Reactions (Exercises)
- Page ID
- 170502

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)P10.1: Draw a mechanism showing the formation of an imine linkage between a lysine side chain and a-ketobutyrate (this is the first step in the degradation of lysine, EC 1.5.1.8).
P10.2: Draw four possible cyclic hemiketal isomers of the compound below.
P10.3: A downstream intermediate in the lysine degradation pathway undergoes imine hydrolysis to release two amino acid products (EC 1.5.1.1). Draw a mechanism for this hydrolysis reaction, and show the structures of the two products formed.
P10.4: Below is the structure of lactose, the sugar found in dairy products.
Lactose is a disaccharide of galactose and glucose. People who are lactose intolerant do not produce enough lactase - the enzyme that hydrolyzes the glycosidic bond linking the two monosaccharides - to be able to fully digest dairy products.
- Draw a likely stabilized carbocation intermediate in the hydrolysis reaction catalyzed by lactase.
- Draw, in the chair conformation, the structure of what you predict would be the most abundant form of the galactose monosaccharide in aqueous solution.
- Is galactose an aldose or a ketose?
- Draw, showing sterochemistry, the open-chain form of galactose.
P10.5: You probably know that ascorbic acid (vitamin C) acts as an antioxidant in the body. When vitamin C does its job, it ends up being oxidized to dehydroascobate, which is usually drawn as shown below, in the so-called tricarbonyl form.
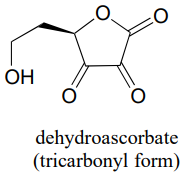
Evidence suggests, however, that the most important form of dehydroascorbate in a physiological context is one in which one of the ketone groups is in its hydrated form, and the other is an intramolecular hemiketal (see Chemical and Engineering News, Aug. 25, 2008, p. 36). Show the structure of this form of dehydroascorbic acid.
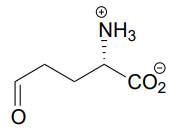
P10.6: The compound below is the product of a ring-opening imine hydrolysis step in the degradation pathway for proline, one of the amino acids. Draw the structure of the starting compound.
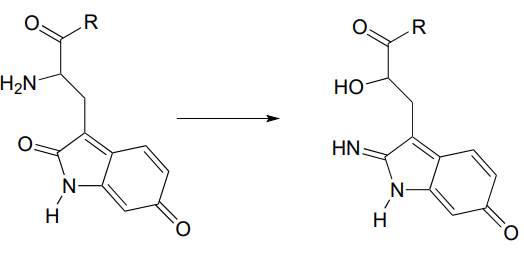
P10.7: The rearrangement below was proposed to proceed via imine formation followed by nucleophilic substitution. Propose a mechanism that fits this description. (J. Biol. Chem. 280, 12858, scheme 2 part 2) .
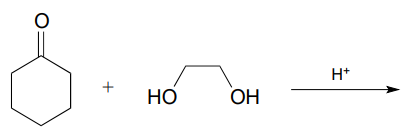
P10.8: The biochemical acetal-forming reactions we learned about in this chapter all require activation of the hemiacetal through phosphorylation. In the organic synthesis lab, non-enzymatic acetal-forming reactions are carried out with a catalytic amount of strong acid, which serves to activate the hemiacetal. Predict the product of the following acetal forming reaction, and propose a reasonable mechanism for the reaction. Remember that the reaction is carried out under acidic conditions, which means that the protonation state of intermediates will be different than biochemical reactions occurring at neutral \(pH\).
Problems 9-15 all involve variations on, and combinations of, the nucleophilic addition steps that we studied in this chapter. Although the reactants and/or products may look somewhat different from the simpler aldehydes, acetals, imines, etc. that we used as examples in the chapter, the key steps still involve essentially the same mechanistic patterns. Before attempting these problems, you may want to review tautomerization reactions in section 7.6.
P10.9: The final step in the biosynthesis of inosine monophosphate (IMP, a precursor to both AMP and GMP), is a ring-closing reaction in which a new nitrogen-carbon bond (indicated by an arrow in the structure below) is formed. Predict the starting substrate for this reaction, and propose a mechanism that involves a slight variation on typical imine formation. (EC 3.5.4.10)
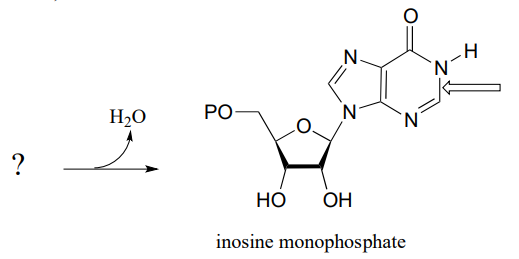
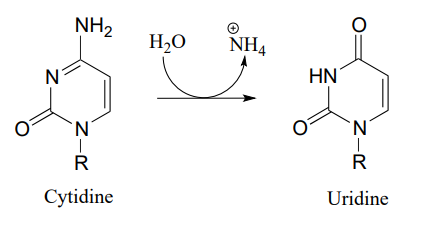
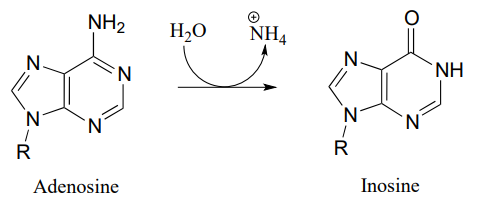
P10.10: Propose a mechanism for these steps in nucleotide metabolism:
- (EC 3.5.4.5)
- (EC 3.5.4.4)
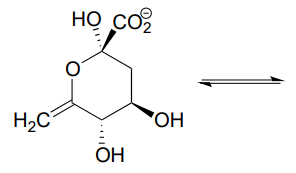
P10.11
- Draw the structure (including stereochemistry) of the compound that results when the cyclic hemiketal shown below coverts to an open-chain compound with two ketone groups.
- The compound shown below undergoes a ring-opening reaction to form a species that can be described as both an enol and an enamine. Draw the structure (including stereochemistry) of this product, and a likely mechanism for its formation. (EC 5.3.1.24)
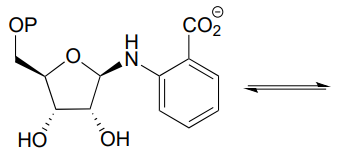
P10.12: Tetrahydrofolate (\(THF\)) is a coenzyme that serves as a single-carbon donor in many biochemical reactions. Unlike \(S\)-adenosylmethionine (SAM, see section 8.8), the carbon being transferred in a \(THF\)-dependent reaction is often part of a carbonyl. Below is a reaction in the histidine degradation pathway (EC 3.5.3.8). The mechanism involved is thought to be an transimination, followed by a imine-to-imine tautomerization, followed by an imine hydrolysis. Propose a reasonable mechanism that fits this description. Hint: first identify the carbon atom being transferred.
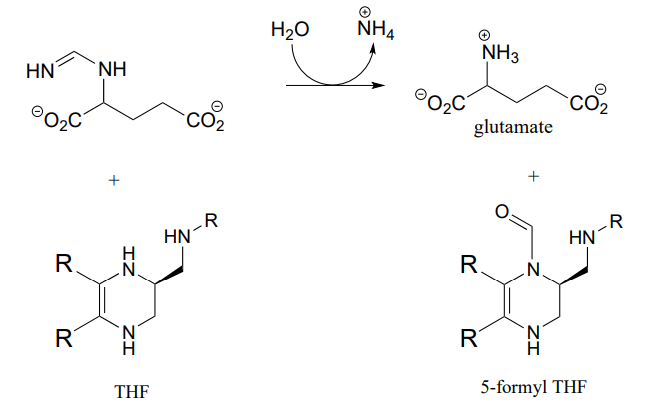
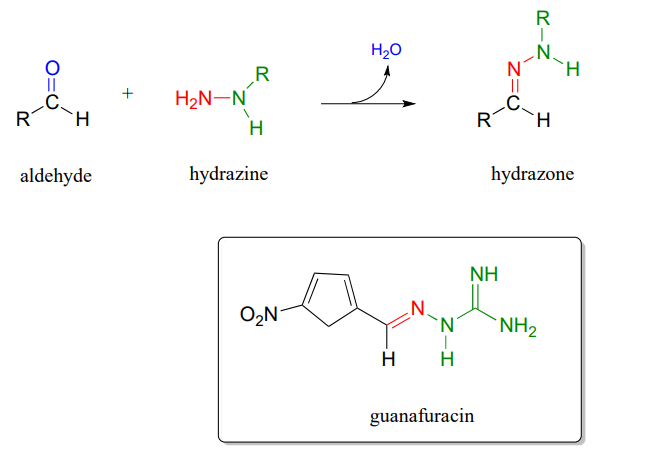
P10.13: Hydrazones are close relatives of imines, formed in reactions between aldehydes/ketones and hydrazines, a functional group containing a nitrogen-nitrogen bond. The mechanism for hydrazone formation is analogous to that of imine formation.
Guanafuracin, a known antibiotic compound, is a hydrazone, and can be prepared easily in the laboratory by combining equimolar amounts of the appropriate aldehyde and hydrazine in water (no heat or acid catalyst is required, and the reaction is complete in seconds).
Determine the starting materials required for the synthesis of guanafuracin, and propose a likely mechanism for the reaction.
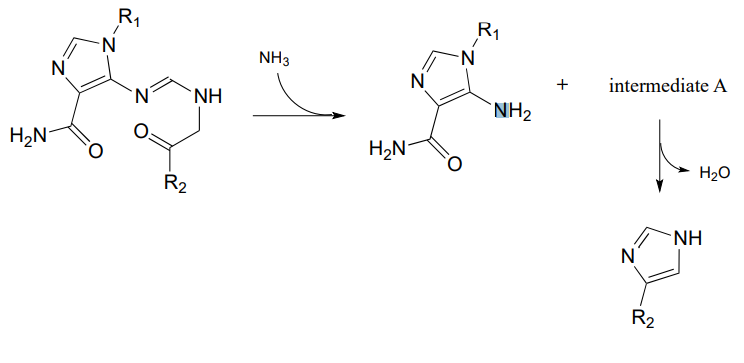
P10.14: Propose reasonable mechanisms for the following steps from the histidine biosynthesis pathway, and predict the structure of intermediate A (which is open-chain, not cyclic).
The last several problems are quite challenging!
P10.15: Propose a likely mechanism for the synthesis of glucosamine 6-phosphate from fructose-6-phosphate. One of several intermediates is shown. (EC 2.6.1.16.)
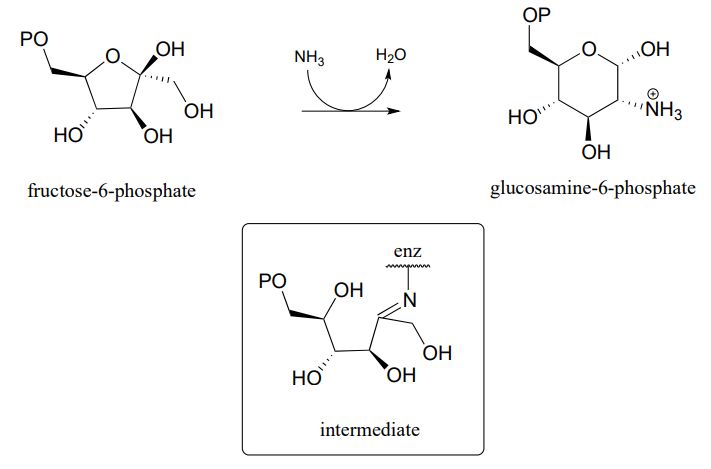
P10.16: \(\alpha \)-chloromethyl ketones (structure below) are effective irreversible inhibitors of proteolytic (peptide-bond breaking) enzymes such as chymotrypsin. In these enzymes, a nucleophilic serine plays a key role in the reaction. The mechanism for inactivation of a-chymotrypsin is thought to involve, as a first step, nucleophilic attack by the active site serine on the carbonyl of the inhibitor. However, when the inactivated enzyme is analyzed, an active site histidine rather than the serine, is found to be covalently modified by the inhibitor. The structure of the modified histidine is shown below. The mechanism of inactivation is thought to involve an epoxide intermediate - with this in mind, propose a reasonable mechanism of inactivation.
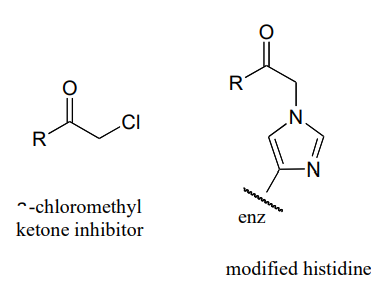
P10.17: An enzyme in E. coli bacteria catalyzes the hydrolysis of a-glucose-GDP to glucose.
\(^1H-NMR\) analysis of the reaction in progress showed the initial appearance of a doublet at 4.64 ppm with J = 7.9 Hz (the spectrum contained other signals as well, of course). After 20 minutes (at which point the hydrolysis reaction has been complete for some time), another doublet began to appear slightly downfield, this one with J = 4.0 Hz. Over time, the strength of the downfield signal gradually increased and that of the upfield signal gradually decreased, until they stabilized at constant levels.
Draw a mechanism for the enzymatic hydrolysis reaction, and correlate your mechanism to the \(NMR\) data (including the appearance of the second doublet).
P10.18: Arginine deaminase, an enzyme in the arginine degradation pathway, catalyzes the transformation of (\(L\))-arginine to (\(L\))-citrulline via a covalent substrate-cysteine intermediate.
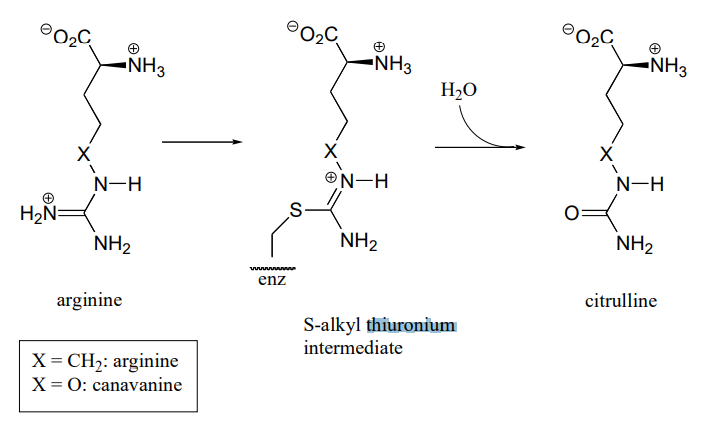
This enzyme is the target for the development of drugs for cancer and immunological diseases such as arthritis. However, rather than completely and permanently shutting down the enzyme (eg. with an irreversible inhibitor), researchers are looking for a way to temporarily 'turn down' the activity of the enzyme. One strategy that has recently been reported involves the use of an oxygen-containing arginine analog, called canavanine, which reacts in the same way as arginine except that the second (hydrolysis) step is very slow. While the enzyme is covalently attached to the inhibitor (in the \(S\)-alkyl thiuronium stage), it is inactivated.
- Show a mechanism for the reaction catalyzed by arginine deaminase.
- Explain how the electronic effect of the oxygen substituent would slow down the hydrolysis step of the reaction, and why the rate of the hydrolysis step is more affected by the oxygen substitution than the \(S\)-alkylthiuronium-forming step.