
12.2: DNA Replication
- Page ID
- 165331

According to the central dogma of molecular genetics, DNA is the genetically active component of the chromosomes of a cell. That is, DNA in the cell nucleus contains all the information necessary to control synthesis of the proteins, enzymes, and other molecules which are needed as that cell grows, carries on metabolism, and eventually reproduces. Thus when a cell divides, its DNA must pass on genetic information to both daughter cells. It must somehow be able to divide into duplicate copies. This process is called replication.
Given the complementary double strands of DNA, it is relatively easy to see how DNA as a molecule is well structured for replication. Each strand serves as a template for a new strand. Thus, after DNA is replicated, each new DNA double helix will have one strand from the original DNA molecule, and one newly synthesized molecule. This is referred to as semiconservative replication.
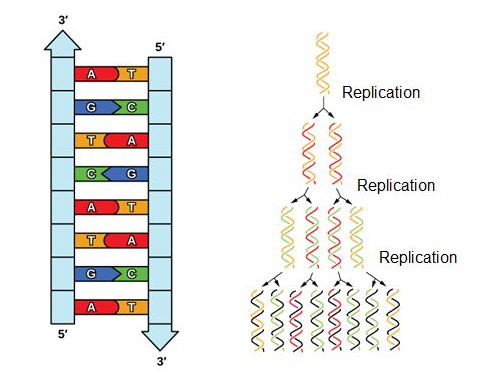
Because eukaryotic genomes are quite complex, DNA replication is a very complicated process that involves several enzymes and other proteins. It occurs in three main stages: initiation, elongation, and termination.
Key Points
- During initiation, proteins bind to the origin of replication while helicase unwinds the DNA helix and two replication forks are formed at the origin of replication.
- During elongation, a primer sequence is added with complementary RNA nucleotides, which are then replaced by DNA nucleotides.
- During elongation the leading strand is made continuously, while the lagging strand is made in pieces called Okazaki fragments.
- During termination, primers are removed and replaced with new DNA nucleotides and the backbone is sealed by DNA ligase
- origin of replication: a particular sequence in a genome at which replication is initiated
- leading strand: the template strand of the DNA double helix that is oriented so that the replication fork moves along it in the 3′ to 5′ direction
- lagging strand: the strand of the template DNA double helix that is oriented so that the replication fork moves along it in a 5′ to 3′ manner
Initiation of Replication
The replication begins at specific nucleotide sequences called origins of replication along the length of the DNA. The circular E. coli chromosome has just one of these sites; the linear eukaryotic chromosomes, in contrast, have multiple sites on every chromosome. Once this site is identified, proteins generally called "initiators" have the capacity to bind DNA at or very near the DNA sequences that mark the origins of replication. The interaction of the initiator proteins with the DNA helps to destabilize the double helix and also to recruit other proteins, including an enzyme called a helicase. In this case the energy required to destabilize the DNA double helix seems to come from the formation of new associations between DNA and the initiator proteins. The DNA helicase, in contrast, once loaded onto the origin, couples the exergonic hydrolysis of ATP to the unwinding of the DNA double helix. Additional proteins must be recruited to the partially unwound initiation complex. These include, but are not limited to, enzymes called primase and DNA polymerase. While the initiators are lost soon after the initiation of replication, the rest of the proteins work in concert to execute the process of DNA replication. This complex of enzymes function at Y-shaped structures in the DNA called replication forks (see figure below). For any replication event two replication forks may be formed at each origin of replication, extending in both directions.
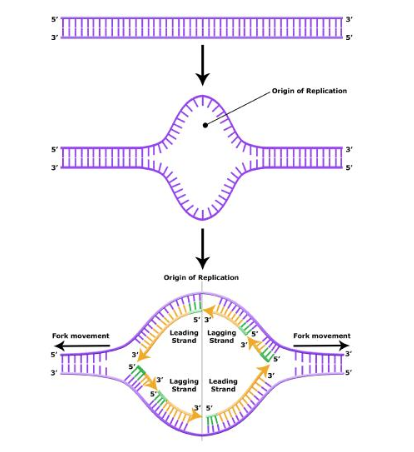
Figure 12.2.2: At the origin of replication, a replication bubble forms. The replication bubble is composed of two replication forks, each traveling in opposite directions along the DNA. The replication forks include all of the enzymes required for replication to occur.
Elongation of Replication
During elongation, an enzyme called DNA polymerase adds DNA nucleotides to the 3′ end of the newly synthesized polynucleotide strand. The template strand specifies which of the four DNA nucleotides (A, T, C, or G) is added at each position along the new chain. Only the nucleotide complementary to the template nucleotide at that position is added to the new strand.
DNA polymerase contains a groove that allows it to bind to a single-stranded template DNA and travel one nucleotide at at time. For example, when DNA polymerase meets an adenosine nucleotide on the template strand, it adds a thymidine to the 3′ end of the newly synthesized strand, and then moves to the next nucleotide on the template strand. This process will continue until the DNA polymerase reaches the end of the template strand.
DNA polymerase cannot initiate new strand synthesis; it only adds new nucleotides at the 3′ end of an existing strand. All newly synthesized polynucleotide strands must be initiated by a specialized RNA polymerase called primase. Primase initiates polynucleotide synthesis and by creating a short RNA polynucleotide strand complementary to template DNA strand. This short stretch of RNA nucleotides is called the primer. Once RNA primer has been synthesized at the template DNA, primase exits, and DNA polymerase extends the new strand with nucleotides complementary to the template DNA.
Eventually, the RNA nucleotides in the primer are removed and replaced with DNA nucleotides. Once DNA replication is finished, the daughter molecules are made entirely of continuous DNA nucleotides, with no RNA portions.
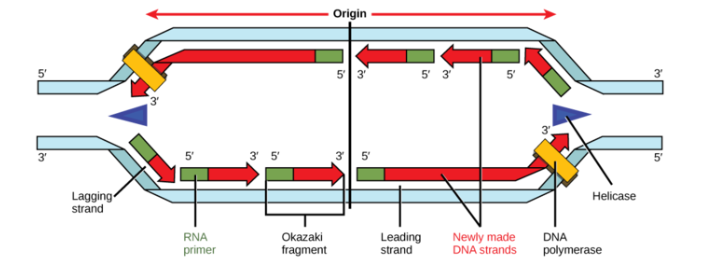
The Leading and Lagging Strands
DNA polymerases can only make DNA in the 5' to 3' direction, and this poses a problem during replication. A DNA double helix is always anti-parallel; in other words, one strand runs in the 5' to 3' direction, while the other runs in the 3' to 5' direction. This makes it necessary for the two new strands, which are also antiparallel to their templates, to be made in slightly different ways.
One new strand, which runs 5' to 3' towards the replication fork, is the easy one. This strand is made continuously, because the DNA polymerase is moving in the same direction as the replication fork. This continuously synthesized strand is called the leading strand.
The other new strand, which runs 5' to 3' away from the fork, is trickier. This strand is made in fragments because, as the fork moves forward, the DNA polymerase (which is moving away from the fork) must come off and reattach on the newly exposed DNA. This tricky strand, which is made in fragments, is called the lagging strand.
The small fragments are called Okazaki fragments, named for the Japanese scientist who discovered them. The leading strand can be extended from one primer alone, whereas the lagging strand needs a new primer for each of the short Okazaki fragments.
Termination of Replication
Eukaryotic chromosomes have multiple origins of replication, which initiate replication almost simultaneously. Each origin of replication forms a bubble of duplicated DNA on either side of the origin of replication. Eventually, the leading strand of one replication bubble reaches the lagging strand of another bubble, and the lagging strand will reach the 5′ end of the previous Okazaki fragment in the same bubble.
DNA polymerase halts when it reaches a section of DNA template that has already been replicated. However, DNA polymerase cannot catalyze the formation of a phosphodiester bond between the two segments of the new DNA strand, and it drops off. These unattached sections of the sugar-phosphate backbone in an otherwise full-replicated DNA strand are called nicks.
Once all the template nucleotides have been replicated, the replication process is not yet over. RNA primers need to be replaced with DNA, and nicks in the sugar-phosphate backbone need to be connected.
The group of cellular enzymes that remove RNA primers include the proteins FEN1 (flap endonulcease 1) and RNase H. The enzymes FEN1 and RNase H remove RNA primers at the start of each leading strand and at the start of each Okazaki fragment, leaving gaps of unreplicated template DNA. Once the primers are removed, a free-floating DNA polymerase lands at the 3′ end of the preceding DNA fragment and extends the DNA over the gap. However, this creates new nicks (unconnected sugar-phosphate backbone).
In the final stage of DNA replication, the enzyme DNA ligase joins the sugar-phosphate backbones at each nick site. After ligase has connected all nicks, the new strand is one long continuous DNA strand, and the daughter DNA molecule is complete.
Figure 12.2.4: Termination of replication
References
- ↑ Mariana Ruiz. Wikimedia Commons. 24 January 2007. Retrieved: 11 August 2009.
- ↑ Nelson, D.L., Cox, M.M. Lehninger Principles of Biochemistry(5thed). New York: W.H. Freeman and Company, 2008. pp. 985-991.
- ↑ Nelson, D.L., Cox, M.M. Lehninger Principles of Biochemistry(5thed). New York: W.H. Freeman and Company, 2008. pp. 982-991.
Contributors
Ed Vitz (Kutztown University), John W. Moore (UW-Madison), Justin Shorb (Hope College), Xavier Prat-Resina (University of Minnesota Rochester), Tim Wendorff, and Adam Hahn.